Fernanda de Mello1*, Juliana Saraiva Garcia1, Alexandra Depince2, Catherine Labbé2 and Danilo Pedro Streit Jr1
1Federal University of Rio Grande do Sul, Department of Animal Science, Porto Alegre 91540-000, RS, Brazil
2INRA, UR1037, Fish Physiology and Genomics, Campus de Beaulieu, 35000 Rennes, France
*Corresponding Author:
Fernanda de Mello
Federal University of Rio Grande do Sul, Department of Animal Science
Porto Alegre 91540-000, RS, Brazil
Tel: +55-51-3308-6853
Fax: +55-51-3308-6048
E-mail: fernandade.mello@gmail.com
Received date: 10.08.2015 / Accepted date: 22.02.2016 / Published online: 28.02.2016
Keywords
Aquaculture; DNA Extraction; Cryopreservation; Epigenetics Alteration; Sperm Cells
Introduction
Tambaqui (Colossoma macropomum) is an important species in South America and the main native species in Brazil commercial production (Ministério da Pesca e Aquicultura 2013). Due to the large consumer market and consequently the great economic importance of this species, industry and government have heavily supported a large contribution to the development of biotechnology for efficient production. Thus, tambaqui has become an important experimental model for reproduction studies, physiology, genetics and cryobiology.
The study on the quality of gametes and semen DNA integrity may answer many questions that guide the optimization of cryopreservation protocols specific-species in fish. However, simple questions to be analyzed become slow to be answered due to troubles in the execution of basic techniques. Currently, many studies are focused on impact of cryoprotectant solutions on the gametes quality, DNA integrity and silencing of important genes for embryonic development (Bobe and Labbé 2010; Valdebenito et al. 2015). To perform mutagenic and epigenetic analysis, DNA isolation is the point of departure, without which is not possible to performer experiments.
Different protocols were described for DNA isolation from semen fish, resulting at good quantity and quality of DNA obtained. Many times, is observed that transfers of DNA extraction protocols among species have not success. However, large parts of advances on knowledge of native fish species are a result of adaptation and / or optimization of protocols described for model species, such as rainbow trout and zebrafish. There is no protocol described for DNA isolation from Tambaqui semen or other species evolutionarily close. Particularly about Tambaqui, semen cryopreservation protocols are extensively optimized which has provided the largely use of cryopreserved genetic material in Tambaqui’s production, species of great economic importance and ecological. Nevertheless, cryoprotectants agents (CPAs) may be a complication factor in DNA extraction because add chemicals that can affect the quality of extracted DNA interfering in subsequent techniques like polymerase chain reaction (PCR) and restriction endonucleases, which require a DNA quality (Langhans and Palladino 2009).
In the present paper, we used an improved protocol developed to fish muscle for isolated DNA of spermatozoa from fresh and cryopreserved semen. The protocol provides high-quality DNA and useful as template in enzymatic reactions experiments and allows the maintenance of the individuals through germplasm banks.
Material and Methods
Semen samples
Tambaqui (C. macropomum) DNA was obtained from fresh and cryopreserved semen by changing improved protocol previously described for DNA extraction from muscle preserved at different fish species (Wasko et al. 2003). Was used semen collected from three males C. macropomum (2.548 kg ± 0.15) shown good nutritional conditions, which showed at least 90% of motility. Freezing solutions were prepared using five CPAs: Dimethylformamide (DMF), dimethylsulfoxide (DMSO), methanol, ethylene glycol and glycerol (all at 10% concentration) and extended in BTS (Beltsville Thawing Solution). Semen with CPAs was diluted in 1:4 ratio (semen:freezing solution) and loaded into 2.5 ml plastic straw, exposed to liquid nitrogen (NL) vapor for 10 min and then plunged into NL. Fresh semen was collected in cryotubes and immediately stocked at the NL (-196 ºC). Semen was thawed after 24 h and the DNA was isolated from both cryopreserved and fresh semen.
DNA Isolation from fresh and cryopreserved semen
Fresh and cryopreservation semen were thawed, transferred to microtubes and centrifuged for 2 minutes at 7.000 g to the spermatozoa were for the tube bottom. Then, 5 μl were recovered (final concentration ≈ 0.25x109 spz/mL) from the bottom of tube and added to 195 μl of Hanks' Balanced Salt Solution (Ca2+ and Mg2+: 1.26 mM CaCl2, 0.49 mM MgCl2.6H2O, 0.41 mM MgSO4.7H2O, 5.3 mM KCl, 0.44 mM KH2PO4, 4.17 mM NaHCO3, 137.93 mM NaCl, 0.34 mM Na2HPO4, 5.55 mM Dextrose), totaling 200 μl. This volume was adjusted to 1 mL with TNES digestion buffer (10 mM Tris-HCl pH 8.0; 125 mM NaCl; 10 mM EDTA pH 8.0; 0.5 %SDS; 4M urea) inside a 2 ml tube. Then, 3.75 μl proteinase K (20 mg/ml) was added and the sampled were maintained at 42°C for overnight with gentle shaking.
Protein precipitation was performed by transfer 1 ml of the digested sample in a 2 ml microtube adding 1 ml of phenol:chloroform:isoamyl alcohol (25:24:1) (InvitrogenTM) to each tube. After shake vigorously, the tubes for 5-10 seconds, the samples remained 15 min at room temperature and then, were centrifuged for 15 min at 8.000 g (4 ºC). This step separates the sample in two phases: a lower organic phase (containing the lipids and other hydrophobic contents) and an upper aqueous phase (containing dissolved DNA). The interphase is proteins. The top aqueous layer was transfer 400 μl of the upper aqueous phase to DNA isolation to a fresh microtube. DNA was precipitated adding 100 μl of 5M NaCl and 1 ml of absolute ethanol (stored at -20°C) and invert to mix (5 times). The samples remained incubate 15 minutes at room temperature and were centrifuged for 15 minute at 8000 g (4°C) to pellet of DNA visualization. The DNA pellet was washed briefly in 100 μl of 70 % ethanol, air dried and resuspended in TE buffer (10 mM Tris HCl pH 8.0; 1 mM EDTA pH 8.0) with 100 μg/ml RNase in H2O. After completely soluble, the DNA was stored at −20°C.
The DNA concentration was checked in NanoDrop (Thermo Fisher Scientific®) and the DNA quality was checked on agarose gel (0.8%).
Restriction enzyme analysis
In order to test the quality of the DNA isolated 1 μg was incubated with 5 units each of MspI and HpaII (New England, Biolabs) in the recommended buffer at 37°C for 3 h. DNA digestion was assayed by visual inspection after agarose gel electrophoresis.
Results and Discussion
We found some difficulties to work with tambaqui’s spermatozoa due to the high concentration of seminal fluid proteins, which can lead to a low amount and poor quality of total recovered DNA. In fish, differently of the mammals where the gametes are release into female genital tract, fish gametes are release in the aquatic environment; thus, spermatozoa and seminal plasma components must to be at high concentration. Seminal proteins are essential to sperm have the ideal conditions respond to environment signals like osmotic pressure, ionic and gaseous components of external media and, in some cases, egg-derived substances (Dzyuba and Cosson 2014). Tests using a final concentration of 0.075 mg/ml of Proteinase K provided lower concentration of DNA after a 10 hours-incubation, probably due to not complete dissociation of spermatozoa from seminal fluid (Weyrich 2012). The better results were obtained leaving the sample overnight at 42 ºC. Lesstime incubation was not efficient.
An improvement on the DNA isolation process was provided by previous centrifugation at lower rotation, which was effective to make the spermatozoa go to the tube bottom. This step enabled to recover more cells with a lower volume of seminal plasma and CPAs, facilitating the process of DNA isolation. Furthermore, previous centrifugation was essential step to increasing and standardizes the number of spermatozoa in the sample. The standardization of cell number (spermatozoa) for applying the extraction protocol is important because it prevents the excess material, which may not be all processed due to concentration of reagents be limited (Hulme et al. 2013). Thus, excess cells will disrupt the initial protocol steps such as membranes rupture and proteins cleavage, compromising the isolation of total DNA.
The use of phenol-chloroform proved to be essential to obtaining pure DNA samples from fresh and cryopreserved semen. Eliminate residues of protein and CPAs were the major preoccupations in DNA isolation, but single and double washes gave greats results (Figure 1). The isolated DNA had no sign of degradation and the absorbance at 260 – 280 nm measured by nanodrop indicated a DNA/RNA relationship of 1.6 – 2.1, indicating a good DNA quality. The DNA concentration ranged from 0.1–2μg, depending on the number of cells initially recovery, being high enough to be employed on several molecular experiments.
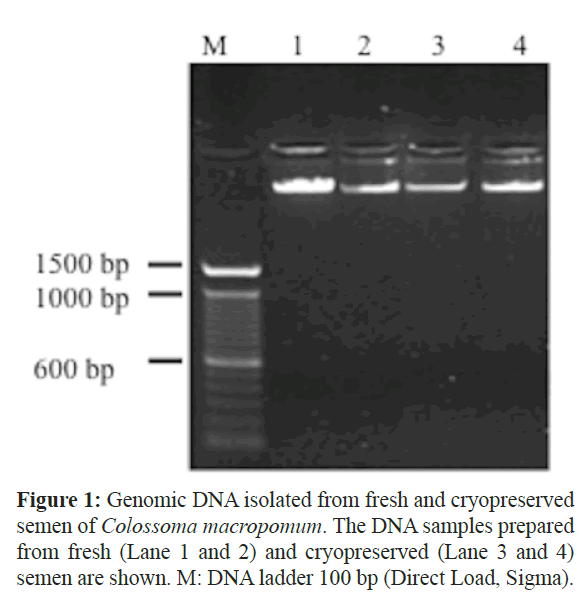
Figure 1: Genomic DNA isolated from fresh and cryopreserved semen of Colossoma macropomum. The DNA samples prepared from fresh (Lane 1 and 2) and cryopreserved (Lane 3 and 4) semen are shown. M: DNA ladder 100 bp (Direct Load, Sigma).
In order to test the quality, isolated DNA was digested with restriction enzymes MspI and HpaII (Figure 2). MspI and HpaII are usually used to epigenetics experiments because they recognize areas amenable to methylation, the CpG islands (Waalwijk and Flavell 1978; Gardiner-Garden and Frommer 1987). These sequences are present in promoter regions of actively transcribed genes and also close to the first exon (Fulnecek and Kovarík 2014). Despite the presence of polysaccharides visible on agarose gel, the portion of DNA that was cleavage (1μg) not showed polysaccharide remains trapped in agarose gel well. The template DNA was completely digested with the enzymes in all the samples, fresh and cryopreserved semen, which showed the isolated DNA was of high quality.
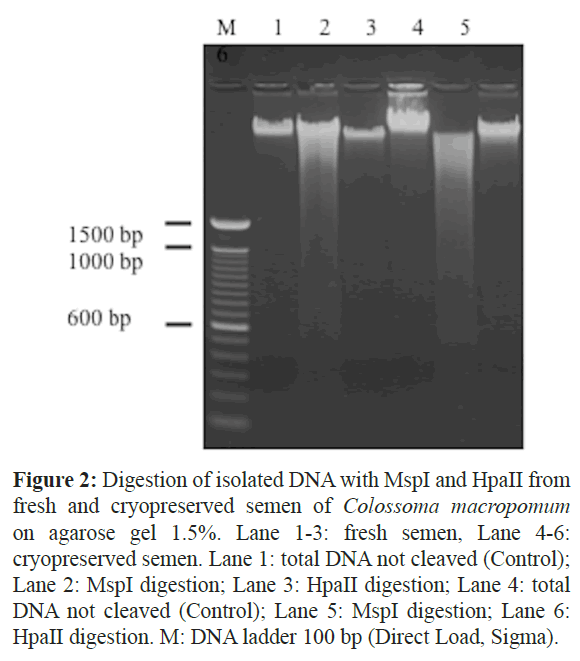
Figure 2: Digestion of isolated DNA with MspI and HpaII from fresh and cryopreserved semen of Colossoma macropomum on agarose gel 1.5%. Lane 1-3: fresh semen, Lane 4-6: cryopreserved semen. Lane 1: total DNA not cleaved (Control); Lane 2: MspI digestion; Lane 3: HpaII digestion; Lane 4: total DNA not cleaved (Control); Lane 5: MspI digestion; Lane 6: HpaII digestion. M: DNA ladder 100 bp (Direct Load, Sigma).
Conclusion
Modern studies on gamete quality increasingly rely on DNA for detecting epigenetics alterations due the cryopreservation and/or exposure to CPAs. The present methodology showed the success in DNA isolation from fresh and cryopreserved semen from different CPAs suitable for use in molecular analyzes.
Acknowledgements
We thank to Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the scholarship granted to the first author.
9182
References
- Bobe, J. and Labbé, C. (2010) Egg and sperm quality in fish.General and comparative endocrinology 165, 535–548
- nDzyuba, V. and Cosson, J. (2014) Motility of fish spermatozoa: from external signaling to flagella response. Reproductive Biology 14, 165–175
- nFulnecek, J. and Kovarík, A. (2014) How to interpret Methylation Sensitive Amplified Polymorphism (MSAP) profiles? BMC Genetics 15, 2
- nGardiner-Garden, M. and Frommer, M. (1987) CpG islands in vertebrate genomes. Journal of Molecular Biology 196, 261–282
- nHulme, P., Lewis, J. and Davidson, G. (2013) Sperm elution: an improved two phase recovery method for sexual assault samples. Science & Justice: Journal of the Forensic Science Society 53, 28–33
- nLanghans, M.T. and Palladino, M.J. (2009) Cleavage of mispairedheteroduplex DNA substrates by numerous restriction enzymes. Current issues in molecular biology 11, 1–12
- nMinistério da Pesca e Aquicultura (2013) Boletimestatístico da pesca e aquicultura: Brasil 2011
- nValdebenito, I.I., Gallegos, P.C. and Effer, B.R. (2015) Gamete quality in fish: evaluation parameters and determining factors. Zygote (Cambridge, England) 23, 177–197
- nWaalwijk, C. and Flavell, R.A. (1978) MspI, an isoschizomer of hpaII which cleaves both unmethylated and methylated hpaII sites. Nucleic Acids Research 5, 3231–3236
- nWasko, A.P., Martins, C., Oliveira, C. and Foresti, F. (2003) Non-destructive genetic sampling in fish.An improved method for DNA extraction from fish fins and scales.Hereditas138, 161–165
- nWeyrich, A. (2012) Preparation of genomic DNA from mammalian sperm. Current Protocols in Molecular Biology / Edited by Frederick M. Ausubel ... [et Al.] Chapter 2, Unit 2.13.1–3.


