Keywords
Feeding behavior; Flatfish; Immunostimulants; Prebiotics; Republic of Korea
Abbreviations: Glucan: Beta-1,3/1,6-glucans; MOS: Mannan Oligosaccharides; South Korea: Republic of Korea; SGR: Specific Growth Rate; HIS: Hepatosomatic Index; FCR: Feed Conversion Ratio; K: Condition Factor; TP: Total Protein Concentration; TCHO: Total Cholesterol Concentration; GLU: Total Glucose Concentration; ALB: Total Albumin Concentration; AST: Total Aspartat-Aminotransferase Concentration; CP: Crude Protein
Introduction
Within the last 30 years the demand for seafood for human consumption has increased globally and is satisfied by expanding aquaculture production while fisheries yields stagnate (FAO, 2014). In the Republic of Korea (South Korea) the farming of flat fish, such as the Olive Flounder (Paralichthys olivaceus, approx. 42,133 t in 2014) (FAO, 2016) has been a common practice for more than 20 years. The aquaculture candidate species P. stellatus is an important fishery resource in South Korea, and has become a target species for prospective aquaculture diversification using production techniques already applied for related pleuronectids like P. olivaceus (An et al., 2011). Its high market value and its adaptive abilities to varying water conditions, e.g. salinity and temperature, make the Starry flounder an interesting candidate also for cultivation in temperate regions (Song et al., 2014, Lee et al., 2003, Ma et al., 2014). However, there is still various knowledge gaps related to the commercial culture of the Starry flounder, such as appropriate formulated diet composition as well as the feeding behaviour.
In the course of continuously intensifying aquaculture production, the demand for higher productivity at lower cost is omnipresent (Rana et al., 2009). One important topic of interest is the reduction of numbers of individuals lost through infections by pathogens. Commercial vaccines are expensive and ineffective against many bacterial diseases and those caused by viruses (Hill, 2005) Additionally, the excessive use of antibiotics can cause a transfer of antibiotic resistance determinants from fish pathogens to human pathogens (Cabello, 2006). As a consequence, in recent years a number of feed additives have been investigated and applied in aquaculture (e.g. prebiotics, probiotics, polysaccharides, nucleotides, acidifiers) (RingØ et al., 2010, Wang et al., 2008, Gatesoupe, 2005, Ganguly et al., 2013, Nakagawa et al., 2007). A positive effect in terms of growth and immune response has been shown for a variety of different substances, such as plant extracts as described by Nakagawa et al. (2007), nucleotides (RingØ et al., 2010, Li and Gatlin Iii, 2006) and oligosaccharides (Grisdale- Helland et al., 2008). Increasing attention has been paid to the use of immunostimulants for disease prevention, especially to those, which enhance non-specific defence mechanisms (Sakai, 1999). Oral administration is favoured for development as it avoids the labour intensive as well as time-consuming method of injection, which was and is commonly applied for juvenile fish. The polysaccharides beta-1,3/1,6-glucans (glucan) and mannan oligosaccharides (MOS), isolated from yeast cell walls or plants have been shown to positively affect growth and health of finfish (Meena et al., 2013, Zeković et al., 2005). Glucan and MOS supplemented feeds demonstrated enhanced growth performance for different carp species, rainbow trout (Oncorhynchus mykiss) and sea bass (Dicentrarchus labrax) (Kühlwein et al., 2014, Misra et al., 2006, Ai et al., 2007, Staykov et al., 2007, Andrews et al., 2009, Torrecillas et al., 2012) but this effect is still not investigated for Starry flounder or other flatfish to date (Lee et al., 2003).
Hence, this study focusses on the use and effect of the feed additives glucan and MOS on the Starry flounder as well as the suitability of Starry flounder as an aquaculture candidate, in terms of health and growth performance. Finally, the feeding behaviour of the flatfish was observed to define its feeding requirements/ suitability under aquaculture conditions.
Materials and Methods
Origin of animals and experimental set-up
Starry flounder juveniles were sourced from a private flatfish hatchery in Gangneung (South Korea). After transfer of the animals to the research facility of the Gangneung-Wonju National University (Marine Research and Education Centre) the fish were kept 2 weeks in a round tank (5,000 L) for acclimation. Prior to the experiment the fish were transferred to the experimental tanks and separated into sub-treatments as outlined in the following.
The experimental setup consisted of 12 tanks of the same design (54 × 34 × 30 cm, 0.18 m2, water level 20 cm ≈ 37 L volume), equipped with adjustable water inlet and outlet pipes and aeration. The entire experimental device was connected to the flow-through system of the research facility. The fresh seawater was pumped from the sea and filtered in sedimentation tanks before it entered the main water storage tank that provided a constant water flow to the experimental sub-devices. The flow-through rate per tank was about 0.6 L*min-1.
Altogether 480 fish with an initial mean body weight of 8.13 (± 2.42) and initial mean standard length of 80.48 mm (± 6.06 mm) were divided into the 12 experimental tanks, resulting in 40 individuals per tank (stocking density 222 fish or 1.81 kg per m²). To ensure equal biomass range per tank, the first 35 fish per tank were picked randomly and weighed and the last 5 fish were picked by weight to adjust the total weight. Six experimental tanks were fed with glucan and MOS enhanced feed, the other half was fed with control feed (Table 1). To minimize tank effects (light input, movement of staff) the treatments were assigned to tanks in an alternating manner. The experiment was carried out for 56 days.
| |
Control |
Glucan/MOS |
| Fishmeal |
55 |
55 |
| Wheat flour |
22 |
21.8 |
| Dehulled soya |
5 |
5 |
| Corn gluten |
5 |
5 |
| Wheat gluten |
5 |
5 |
| Squid liver oil |
5 |
5 |
| Vitamin premix |
1.5 |
1.5 |
| Mineral premix |
1.5 |
1.5 |
| Glucan/MOS |
0 |
0.2 |
| Total |
100 |
100 |
| Nutrient composition [g kg-1] |
| Moisture |
15.28 ± 0.01 |
14.96 ± 0.11 |
| Crude protein |
46.33 ± 0.53 |
45.55 ± 0.06 |
| Crude lipid |
8.01 ± 0.14 |
8.12 ± 0.11 |
| Crude ash |
9.39 ± 0.25 |
10.83 ± 0.30 |
Table 1: Ingredients and nutrient composition in g kg−1 dry matter (DM) of the experimental diets.
Water quality parameters, such as pH, dissolved oxygen, salinity, and temperature were measured on a daily basis for the whole experimental duration. The pH was measured using a SG2–pH meter (SevenGo™, Mettler-Toledo International Inc., Seoul, South Korea); dissolved oxygen, salinity and temperature were measured with a YSI 85 multi-parameter instrument (YSI, Kawasaki City, Japan). Mortalities were recorded daily and were removed from the culture units after documentation.
Feed and feeding regime
All applied feeds were produced at Gangneung-Wonju National University (Marine Research and Education Centre) prior to the experiment. The feed ingredients (Table 1) were mixed in a vertical bakery mixer (HDM Co. Ltd., Seoul, South Korea) and processed in a meat mincer (Hankook Fujee Industries Co. Ltd., Seoul, South Korea) (perforated disc hole Ø=3 mm). The feed cords were broken into pieces of 1-10 mm in length, airdried at room temperature and stored in containers at -20°C. The composition of the control-feed and the glucan/MOS-feed was identical except for the addition of 0.2% glucan/MOS replacing an equal amount of wheat flour. The applied glucan/MOS was an extract from the cell walls of the yeast Saccharomyces cerevisiae and provided as dried powder (ProEn Protein und Energie GmbH, Soltau, Germany).
The fish were fed by hand in constant periodic intervals ad libitum twice a day at 10:00 am and 4:30 pm. The feed used was weighed daily to determine total amount of food per tank per day. During the first two weeks of the experiment a ranking list containing different classifications concerning the feeding behaviour was established evaluating the observed results from 0 to 5, whereas 0 represented no reaction to feed and 5 was a strong feeding response (Table 2). Five relevant factors of feeding behavior for flatfish culture (Mas-Muñoz et al., 2011) were identified prior to the experiments and monitored in the first two weeks of the experiment. These factors are (1) interest in food, (2), food acceptance, (3) up-swimming during feeding, (4) anticipation of the regular feeding intervals, and (5) aggression toward other individuals.
| Experiment day |
| |
1 |
2 |
3 |
4 |
5 |
6 |
7 |
8 |
9 |
10 |
11 |
12 |
13 |
14 |
| Interest in food |
0 |
0 |
1 |
1 |
2 |
3 |
3 |
4 |
5 |
5 |
5 |
5 |
5 |
5 |
| Up swimming during feeding |
0 |
0 |
0 |
1 |
1 |
1 |
1 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
| Anticipation of feeding intervals |
0 |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
2 |
3 |
4 |
4 |
4 |
4 |
| Aggression against specimen |
0 |
0 |
0 |
0 |
1 |
1 |
1 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
| Indication of saturation |
0 |
0 |
0 |
0 |
0 |
1 |
1 |
2 |
2 |
2 |
2 |
2 |
2 |
2 |
Table 2: Results of the feeding behaviour monitoring, classifications concerning the feeding behaviour graded from 0 to 5, whereas 0 represents no reaction and 5 was a strong response, the levels of grey also show the strength of response.
Growth
The Specific growth rate (SGR) was calculated using the formula
SGR (% body weight day-1)=[ln(final weight)-ln(initial weight)]/ feeding days × 100 (1)
The hepatosomatic index (HSI) was calculated according to the formula
HSI (%)=(liver weight/ final body weight) × 100 (2)
where WL is the wet weight of the liver [mg] and WT is the total wet body weight. The feed conversion ratio (FCR) was calculated according to the formula
FCR=feed intake (g)/ weight gain (g) (3)
The condition factor (K) was calculated according to the formula
K (%)=100 × (W × L-3) (4)
Where W is the whole body wet weight in grams and L is the length in centimeters.
Sampling procedure and analysis
Each week, 15 fish per treatment were sampled to obtain samples for further analyses of the fitness of the cultured fish. The individuals were randomly netted and bathed in an immersion with an overdose (500 mg × l-1) of tricaine methane sulfonate (MS 222; Sigma-Aldrich, St. Louis, USA) till death. The fish were removed from solution, carefully dried with a paper towel and weighed. Blood was taken from the caudal aorta with a heparintreated syringe. Afterwards, fish were dissected and the liver, the stomach, and the gut were weighed. All samples were stored in 1.5 ml micro tubes (Eppendorf AG, Hamburg, Germany) at -80°C until further analysis. On day 49, the sampling was not completed due to technical problems with water supply, preliminary results until this point did not show any trends in weekly changes.
The plasma concentration of the blood biochemical parameters total glucose (GLU), total cholesterol (TCHO), albumin (ALB) and glutamate-oxaloacetate-transaminase/aspartate-aminotransferase (GOT/AST) was analyzed using a high-performance drychemistry analyzer (Fuji Dri-Chem 4000) (FUJIFILM Europe GmbH, Düsseldorf, Germany) following the manual instructions. To get a sufficient sample volume for the analyzer, two samples of fish from the same tank were pooled by mixing 60 μl of each sample, respectively. The protein content of the blood plasma was measured following the Bradford Method following the Sigma- Aldrich assay protocol using a Spectrophotometer (Multiscan Spectrum, Thermo Fisher Scientific Inc., Waltham USA). The lysozyme activity was examined using a suspension of 200 mg*l-1 freeze dried Micrococcus lysodeikticus (Ml) in 0.05M Phosphatebuffered saline (chemicals: Sigma-Aldrich, St. Louis, USA) as substrate. The reduction of absorbance at 530 nm displays the relative lysozyme activity in units*min-1. The absorbance was measured immediately and after 20 min incubation at 20°C.
The total amount of carbon and nitrogen in feed and feces was measured using a EURO EA elemental analyzer (HEKAtech GmbH, Wegberg, Germany). The crude protein content (CP) was calculated using the formula
CP=N*6.25
Where N is the nitrogen concentration [g*kg-1] and 6.25 is the nitrogen to protein conversion factor.
The composition of feed and fish was analyzed at the Gangneung-Wonju National University according to the FAO standard analysis (FAO, 1987). Crude lipid content was analyzed by ether extraction method, moisture content with a dry oven (105°C for 24 h), and ash content by a furnace muffler (550°C for 4 h). Crude fiber content was analyzed by autoanalyzer (Fibertec; Foss Tecator AB, Höganäs, Sweden).
For the analysis of the digestive enzymes the intestine samples were homogenized in distilled water, filled in microtubes, centrifuged and the supernatant enzyme solution was used for further analysis. All enzyme activities were analyzed using a Spectrophotometer (Multiscan Spectrum, Thermo Fisher Scientific Inc., Waltham USA). The activity of Trypsin-like serine proteases was measured at 366 nm using azocasein (0.5% Azocasein in 0.1 M Phosphate buffer (pH8); Sigma-Aldrich, St. Louis, USA) as a substrate. The Amylase activity was measured at 540 nm using starch (1% starch in 20 mM sodium phosphate buffer; Sigma- Aldrich, St. Louis, USA) as a substrate. For the Lipase assay paranitrophenyl palmitate (1 ml 10 mM pNPP in 4 ml EtOH; Sigma- Aldrich, St. Louis, USA) was used as a substrate and the activity was measured at 405 nm.
To determine the RNA/DNA-ratio samples of muscle tissue were freeze-dried for 24 hours at -51°C in vacuum (33 Pa) (EYELA-FDU-1200) and homogenized. The RNA/DNA-ratio was measured using a spectral scanning multimode reader (Thermo Scientific Varioskan Flash applying the Thermo Scientific SkanIt 2.4.3 software).
Statistical analysis and statistical modelling
Statistical software sigma plot 12.5 for Windows (Systat Software Inc., San Jose, CA, USA) was used for the statistical analysis. All data were tested for normal distribution and homogeneity of variance before further analysis. In order to determine the significance of differences between treatments a one-way analysis of variance (ANOVA) was applied, followed by the post hoc Tukey’s Honestly Significant Difference (HSD) test if variances of the treatments were homogeneous. In case of non-homogeneous variances, the Dunnett T3 test was applied. All statistical tests were performed for P<0.05. Results are expressed as mean ± standard deviation.
The modeling of weight data was performed using R Version 2.12.0. Time dependency of measured weight of five fish and the effect of glucan/MOS was modeled using log-transformed weight data in a linear model, including linear and quadratic time to ensure that weekly alternating sampling methods did not influence the data. Additionally, a generalized linear model (GLM) from the quasi-Poisson family with logarithm as link function was constructed to compare the total weight dependency, i.e., linear growth rate between treatments.
Results
Culture conditions
Salinity ranged between 33 PSU and 35 PSU, the average temperature 15.32°C (± 2.52°C) decreased from about 19°C (18.85°C ± 0.5°C) in the first two weeks to below 14°C (13.66°C ± 1.35°C) on average in the last two weeks of the experiment period. The oxygen level ranged between 7 mg/L and 8 mg/L. The pH value rose over the experiment period from 8.2 to 8.6. During the whole experiment two fish of the glucan/MOS treatment and three of the control treatment died, resulting in a mortality rate of 0.4% and 0.6% for the glucan/MOS treatment and the control treatment, respectively.
Feeding behavior
The fish accepted the feed from first feeding onwards. During the first three days of the investigation the fish left the ground only for a few centimeters and fed restrictively on sunken pellets. Thereafter, fish fed in the lower half of the water column and on the tank bottom. No adaption to frequent feeding intervals was observed. Signs of saturation were not obvious; the individuals also fed on feed particles on the ground for at least 15 to 30 minutes after feeding. Minimal aggression like biting the fins of other individuals was observed (Table 2). Over the subsequent 14 days feeding behaviour appeared to adapt, with increased levels of all response types, stabilizing at day 11.
Growth performance
Mean final weight and weight gain of sampled animals in the glucan/MOS treatment (30.15 g ± 7.00 g) was not significantly higher than that of the control (26.27 g ± 9.59 g) at the end of the experimental stage (Table 3). The specific growth rate of the glucan/MOS treatment (2.33 ± 0.15 %*day-1) was not significantly higher than that of the control treatment (2.07 ± 0.38% *day-1). The daily feed intake (% BW d-1) of the glucan/MOS treatment (1.17 ± 0.12) was significantly higher (P=0.020) than the control (1.02 ± 0.06). FCR of both treatments did not differ significantly between glucan/MOS (0.66 ± 0.06) and control treatments (0.82 ± 0.28). The hepatosomatic index (HSI) was identical for both treatments with 2.06 (± 0.45%) for the control group and 2.06 (± 0.38) for the glucan/MOS group. The condition factor (K) was 1.69 (± 0.20) for the control and 1.80 (± 0.14) showing no significant difference. Mortality was 0.6% for glucan/MOS treatment and 0.4% for the control treatment.
| |
Control |
GM |
| Initial weight (g) |
8.13 ± 2.42 |
8.13 ± 2.42 |
| Final weight (g) |
26.27 ± 9.59 |
30.15 ± 7.00 |
| Weight gain (g) |
18.14 ± 5.23 |
22.14 ± 2.12 |
| SGR (% day-1) |
2.09 ± 0.38 |
2.33 ± 0.15 |
| DFI (% day-1) |
1.17 ± 0.12 |
1.02 ± 0.06 |
| FCR |
0.82 ± 0.28 |
0.66 ± 0.06 |
| Mortality (%) |
0.4 |
0.6 |
| K (%) |
1.69 ± 0.20 |
1.80 ± 0.14 |
| HSI (%) |
2.06 ± 0.45 |
2.06 ± 0.38 |
| |
|
|
| |
SGR=Specific growth rate, DFI=Daily feed intake, FCR=Feed conversion ratio, K=Condition factor, HSI=Hepatosomatic index. Each value is mean ± S.D. (n=6) or (n=3). No significant differences were identified (p>0.05).
Table 3: Growth performance and feed utilization of starry flounder fed experimental diet for 56 days. Each value is mean ± SD (n=6). No significant differences were identified (p>0.05).
The linear model of log-transformed weight data, showed that weekly alternating sampling from two treatments of three basins had no significant effect (p<0.05) and the quadratic term reported non-significant.
The selected model (GLM) kept linear time effects and group treatment alone, showing that Glucan/MOS treated fish exhibited a significantly higher mean growth rate in Glucan/ MOS vs. control (15%, higher coefficient, p=0.0060) (Figure 1). Additionally a generalized linear model from the quasi-Poisson family with logarithm as link function showed similar results for the total weight dependency, i.e., linear growth rate. Here, model selection ended in effects (13%, p=0.0075, over dispersion 1.63) comparable to the results of linear model.
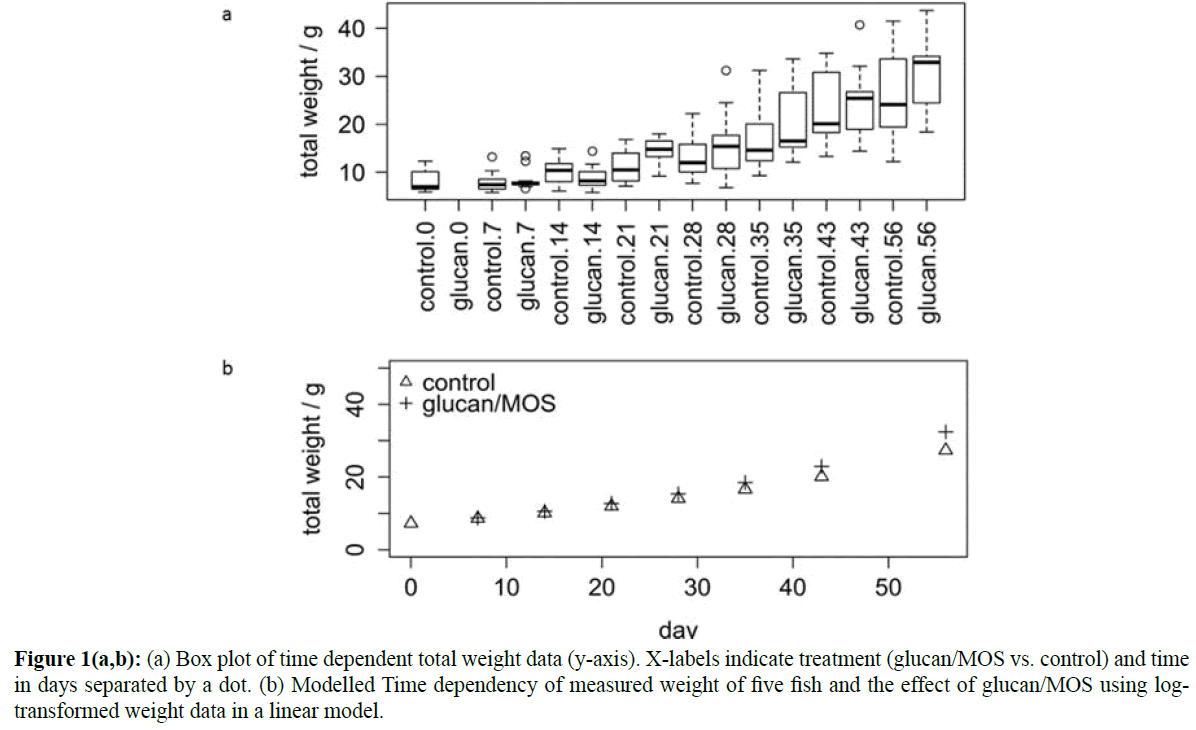
Figure 1(a,b): (a) Box plot of time dependent total weight data (y-axis). X-labels indicate treatment (glucan/MOS vs. control) and time in days separated by a dot. (b) Modelled Time dependency of measured weight of five fish and the effect of glucan/MOS using logtransformed weight data in a linear model.
Health Indicators
The concentrations of all blood parameters were higher for total protein, cholesterol, glucose, albumin and Aspartat Aminotransferase (AST) for the glucan/MOS treatment, significantly for cholesterol (P=0.043) and albumin (P=0.016) (Table 4).
| |
Control |
Glucan/MOS |
| TP (g/dl) |
3.15 ± 0.29 |
3.48 ± 0.39 |
| TCHO (mg/dl) |
220.33 ± 17.39a |
257.33 ± 35.07b |
| GLU (mg/dl) |
43 ± 8.81 |
49 ± 5.97 |
| ALB (g/dl) |
0.85 ± 0.08a |
0.98 ± 0.08b |
| AST (U/I) |
93.17 ± 54.26 |
105.83 ± 69.67 |
| Lysozyme activity (U/min-1) |
25.98 ± 8.09 |
29.03 ± 10.65 |
| RNA/DNA ratio |
3.72 ± 0.84 |
4.22 ± 0.71 |
| Enzyme activity (mU/g tissue) |
| Trypsin |
68658.66 ± 4383.84 |
66401.91 ± 9612.49 |
| Lipase |
3484.04 ± 743.59 |
3130.69 ± 409.79 |
| Amylase |
1598.89 ± 848.23 |
1364.05 ± 122.17 |
| Proximate analysis |
| Moisture |
76.11 ± 0 .51 |
75.35 ± 0.36 |
| Crude protein |
14.49 ± 0.19a |
16.17 ± 0.15 b |
| Crude lipid |
3.73 ± 0.05a |
4.13 ± 0.10 b |
| Ash |
3.18 ± 0.245 |
3.22 ± 0.15 |
| |
|
|
TP=Total protein concentration. TCHO=Total cholesterol concentration. GLU=Total glucose concentration. ALB=Total albumin concentration. AST=Total Aspartat-Aminotransferase concentration. Different superscript letters within a line denote significant differences (p<0.05).
Table 4: Health related parameters: Concentration of blood plasma parameters; Lysozyme activity in plasma; RNA/DNA ratio in muscle tissue, digestive enzyme activity in the intestine and proximate composition of fish and feed: Moisture, crude protein, crude lipid, and ash contents; the values show no significant differences; Mean values ± SD. The values show no significant differences.
Lysozyme activity was higher in fish fed the glucan/MOS diet 29.03 (± 10.65) compared to the control 25.98 (± 8.09) but values were not significantly different. RNA/DNA–ratio was higher in fish fed the glucan/MOS diet than the control diet but the values did not differ significantly. The digestive enzyme activity in the intestine of trypsin, lipase and amylase showed no significant differences between the two treatments. The whole body proximate analysis shows significantly elevated crude protein 16.17% (± 0.15%) (P=<0.001) and lipid 4.13 (± 0.10) (P=<0.005) values for the glucan/MOS fed fish compared to the control group with 14.49% (± 0.19%) and 3.73% (± 0.05%), respectively. Moisture and ash contents were not affected (Table 4). The administered glucan/MOS had no significant effect on the C/N ration of the feces. The protein content of the feces from Starry flounder ranged from approximately 20% to 30%.
Discussion
The development of high value novel aquaculture candidates for continued aquaculture expansion, along with the development of suitable diet additives to sustainably support increased productivity, are key challenges for aquaculture in South Korea and worldwide. In the current study the value of the flatfish Starry flounder, Platichthys stellatus, as an aquaculture species was clearly indicated by robust growth and survival in tank-based feeding experiments. Commensurately, the positive effect of the prebiotic diet additive glucan/MOS on the growth and fitness of P. stellatus was clearly exhibited.
Starry flounder: An aquaculture candidate
Complete culturing of P. stellatus including reproduction control and captive spawning, hatching, and larval and juvenile rearing are possible and practiced in East Asian countries (An et al., 2011). The growth results clearly show the strong early growth rates and excellent feed conversion rate (FCR) values for the species at this phase and underline its potential as a viable aquaculture species. With 2.33%*day-1 (± 0.15%) the growth rate of the glucan/MOS group for this experiment was very good compared to growth rates for starry flounder in other studies <2.4%*day-1 (Wang et al., 2016), <2%*day-1 (Ding et al., 2010), or <1.6 %*day-1, especially taking into account the low temperature during experiment period.
Imsland et al. (2001) could show growth rates for turbot at the same life stage >3% and Choi et al. (2004) showed comparable values for Olive flounder under optimum conditions. But recent studies show lower SGRs for turbot <1.7% (Kroeckel et al., 2012a), <1.8% (van Bussel et al., 2013), <1.5% (Peng et al., 2013), <2.3% (Peng et al., 2014), and for Olive flounder <2% (Pham et al., 2014) <2.7% (Kim et al., 2014) at similar life stages. Considering the suboptimal temperature regime the growth rate achieved in the current study appears impressive. The FCR for the glucan/MOS group (0.66 ± 0.06) was lower but in comparable range to values from Ding et al. (2010) (>0.75) and Wang et al. (2016) (>0.71) for starry flounder and to recent values for turbot >0.76 (Kroeckel et al., 2012b) >1.07 (van Bussel et al., 2012) or Olive flounder >0.86 (Kim et al., 2010). Optimum rearing temperature reported for turbot ranges between 18.3–21.8°C (Cho et al., 2005) and around 20°C for Olive flounder (Okorie et al., 2013). If mentioned, average water temperature in above cited studies was always above 16°C. In this study water temperature (15.32 ± 2.52°C) decreased constantly during the experimental period with fluctuations of more than 1°C per day to less than 14 °C (13.66°C ± 1.35°C) in the last two weeks. Under these conditions growth rate, FCR and feed uptake are especially remarkable. These results suggest that the production of starry flounder in flow through production systems may be favorable in spring and autumn compared to more susceptible species like turbot Olive flounder and can help saving costs in terms of heating and insulation for RAS facilities.
Understanding feeding behavior is essential to adjusting food supply without creating excessive waste (Lagardère and Mallekh, 2000, Mallekh et al., 1998). Feed design and consequent feeding behavior is a crucial factor in animal welfare (Ashley, 2007). In contrast to turbot or Olive flounder starry flounders in this study were more reluctant and did not feed at the water surface. This behavior makes it more difficult to observe feed uptake and satiation but P. stellatus feeding on sunken pellets may improve feed efficiency, although there is dearth of literature to address this topic.
Glucan/MOS Effect
Growth performance results and modelling show a clear growth advantage of glucan/MOS diet inclusion for this species at this growth phase. The immune stimulatory effects of β-glucans on fish have been extensively investigated for a number of fish species with a wide range of results. The treatment enhanced survival rate in challenge tests (Sakai, 1999) and growth performance in a similar way to that observed herein for a number of species (Misra et al., 2006, Ai et al., 2007, Staykov et al., 2007, Yoo et al., 2007, Li et al., 2008, Andrews et al., 2009, Torrecillas et al., 2012, Kühlwein et al., 2014). Yoo et al. (2007) observed a positive effect on weight gain and SGR for Olive flounder (Paralichthys olivaceus), which was fed diets containing yeast glucan for 7 weeks. Fish had a comparable initial weight (9.2 g) but growth performance was highest at 1 and 1.5 g/kg glucan inclusion, which is higher than the applied concentration of 2 g kg-1 GM (20% β-glucan/ 17% MOS) in this study.
The positive effect of glucan/MOS on fish health is supported by significantly enhanced blood parameters albumin (P=0.016) and cholesterol concentration (P=0.043). Maita (2007) stresses that lower cholesterol concentrations increase infection risks and (Andreeva, 2010) describes elevated albumin concentrations in active growth phases of fish. All blood plasma parameters are comparable to those considered as values for fish with sufficient nutrition (Maita, 2007).
The whole body proximate analysis results are comparable to values for Olive flounder (Choi et al., 2004). The significantly higher values for protein and lipid content support the elevated blood parameters in showing the growth rate enhancing effect of glucan/MOS.
The feeding corresponds to standards in literature and the specific growth rate (SGR) shows ranges as suggested by Wang et al. (2015). Bagni et al. (2005) link lysozyme activity and temperature changes. Thus, the high variability of lysozyme activities may be explained by the reduction and variation of water temperature at the end of the experimental period.
The recorded protein content of the feces was relatively high (20 to 30%) compared to values for European seabass (Dicentrarchus labrax) and sea bream (Sparus aurata) (12 to 21%) (Bischoff, 2007), indicating that the administered feed was not completely digested. Starry flounder may have lower protein requirements than D. labrax and S. aurata, allowing the reduction of protein levels in the diet. Analysis of digestive enzymes did not show any differences between the two treatments; hence it can be assumed that the glucan/MOS addition had no negative impact on the activity of the major enzymes in stomach and intestine.
Overall, commercially comparable growth performance, robust health and survivorship in tank holding, and temperature tolerance make the Starry flounder a strong candidate for increasingly intensive aquaculture. The diet additive beta-1-3- glucan and mannans significantly improved growth performance of juvenile starry flounders in a flow through rearing system and associated health parameters like whole body proximate composition and blood plasma parameters support the conclusion that this additive significantly improved the physiological status of treated P. stellatus in the current system.
Acknowledgements
This study was financially supported by NIFS (R2016018), Korea. The first author would like to thank the German Academic Exchange Service (DAAD) for supporting the stay abroad with a travel grant and the Gangneung-Wonju National University (Marine Research and Education Centre) for the great support.
19538
References
- Ai, Q., Mai, K., Zhang, L., Tan, B., Zhang, W., et al. (2007) Effects of dietary β-1, 3 glucan on innate immune response of large yellow croaker, Pseudosciaena crocea. Fish Shellfish Immunol 22, 394-402.
- An, H.S., Byun, S.G., Kim, Y.C., Lee, J.W., Myeong, J.I. (2011) Wild and Hatchery Populations of Korean Starry Flounder (Platichthys stellatus) Compared Using Microsatellite DNA Markers. Int J Mol Sci 12, 9189.
- Andreeva, A.M. (2010) Structure of fish serum albumins. J Evol Biochem Physiol 46, 135-144.
- Andrews, S.R., Sahu, N.P., Pal, A.K., Kumar, S. (2009) Haematological modulation and growth of Labeo rohita fingerlings: effect of dietary mannan oligosaccharide, yeast extract, protein hydrolysate and chlorella. Aquacult Res 41, 61-69.
- Ashley, P.J. (2007) Fish welfare: Current issues in aquaculture. Applied Animal Behaviour Sci 104, 199-235.
- Bagni, M., Romano, N., Finoia, M.G., Abelli, L., Scapigliati, G. et al. (2005) Short- and long-term effects of a dietary yeast β-glucan (Macrogard) and alginic acid (Ergosan) preparation on immune response in sea bass (Dicentrarchus labrax). Fish Shellfish Immunol 18, 311-325.
- Cabello, F.C. (2006) Heavy use of prophylactic antibiotics in aquaculture: a growing problem for human and animal health and for the environment. Environmental Microbiol 8, 1137-1144.
- Cho, S.H., Lee, S.M., Lee, S.M., Lee, J.H. (2005) Effect of dietary protein and lipid levels on growth and body composition of juvenile turbot (Scophthalmus maximus L) reared under optimum salinity and temperature conditions. Aquaculture Nutrition 11, 235-240.
- Choi, S.M., Wang, X., Park, G.J., Lim, S.R., Kim, K.W. et al. (2004) Dietary dehulled soybean meal as a replacement for fish meal in fingerling and growing olive flounder Paralichthys olivaceus (Temminck et Schlegel). Aquacult Res 35, 410-418.
- Ding, L., Zhang, L., Wang, J., Ma, J., Meng, X. et al. (2010) Effect of dietary lipid level on the growth performance, feed utilization, body composition and blood chemistry of juvenile starry flounder (Platichthys stellatus). Aquaculture Res 41, 1470-1478.
- Food and Agriculture Organisation of the United Nations (FAO) (2014) The State of World Fisheries and Aquaculture, Rome.
- Food and Agriculture Organization of the United Nations (FAO) (2016) Fisheries Global Information System (FAO-FIGIS) -Web site. On-going developments-FIGIS, FI Institutional Websites.
- Ganguly, S., Dora, K., Sarkar, S., Chowdhury, S. (2013) Supplementation of prebiotics in fish feed: a review. Rev Fish Biol Fisher 23, 195-199.
- Gatesoupe, J. (2005) Probiotics and prebiotics for fish culture, at the parting of the ways. Aqua Feeds: Formulation & Beyond (Feedware) 2, 3-5.
- Grisdale-Helland, B., Helland, S.J., Gatlin Iii, D.M. (2008) The effects of dietary supplementation with mannanoligosaccharide, fructooligosaccharide or galactooligosaccharide on the growth and feed utilization of Atlantic salmon (Salmo salar). Aquacult 283, 163-167.
- Hill, B.J. (2005) The need for effective disease control in international aquaculture. Developments in biological 121, 3-12.
- Imsland, A.K., Foss, A., Gunnarsson, S., Berntssen, M.H.G., FitzGerald, R., et al. (2001) The interaction of temperature and salinity on growth and food conversion in juvenile turbot (Scophthalmus maximus). Aquacult 198, 353-367.
- Kim, H.S., Jung, W.-G., Myung, S.H., Cho, S.H., Kim, D.S. (2014) Substitution effects of fishmeal with tuna byproduct meal in the diet on growth, body composition, plasma chemistry and amino acid profiles of juvenile olive flounder (Paralichthys olivaceus). Aquacult 431, 92-98.
- Kim, S.S., Pham, M.A., Kim, K.W., Son, M.H., Lee, K.J. (2010) Effects of microbial fermentation of soybean on growth performances, phosphorus availability, and antioxidant activity in diets for juvenile olive flounder (Paralichthys olivaceus). Food Science and Biotechnol 19, 1605-1610.
- Kroeckel, S., Harjes, A.G.E., Roth, I., Katz, H., Wuertz, S., et al. (2012a) When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute - Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquacult 364, 345-352.
- Kroeckel, S., Harjes, A.G.E., Roth, I., Katz, H., Wuertz, S., et al. (2012b) When a turbot catches a fly: Evaluation of a pre-pupae meal of the Black Soldier Fly (Hermetia illucens) as fish meal substitute-Growth performance and chitin degradation in juvenile turbot (Psetta maxima). Aquacult pp: 345-352.
- Kühlwein, H., Merrifield, D.L., Rawling, M.D., Foey, A.D., Davies, S.J. (2014) Effects of dietary β-(1,3)(1,6)-D-glucan supplementation on growth performance, intestinal morphology and haemato-immunological profile of mirror carp (Cyprinus carpio L.). J Animal Physiol and Animal Nutrition 98, 279-289.
- Lagardère, J.P., Mallekh, R. (2000) Feeding sounds of turbot (Scophthalmus maximus) and their potential use in the control of food supply in aquaculture: I. Spectrum analysis of the feeding sounds. Aquacult 189, 251-258.
- Lee, S.M., Lee, J.H., Kim, K.D. (2003) Effect of dietary essential fatty acids on growth, body composition and blood chemistry of juvenile starry flounder (Platichthys stellatus). Aquacult 225, 269-281.
- Li, P., Gatlin Iii, D.M. (2006) Nucleotide nutrition in fish: Current knowledge and future applications. Aquacult 251, 141-152.
- Li, Y., Wang, Y.J., Wang, L., Jiang, K.Y. (2008) Influence of several non-nutrient additives on nonspecific immunity and growth of juvenile turbot, Scophthalmus maximus L. Aquacult Nutrition 14, 387-395.
- Ma, J., Wang, J., Zhang, D., Hao, T., Sun, J., et al. (2014) Estimation of optimum docosahexaenoic to eicosapentaenoic acid ratio (DHA/EPA) for juvenile starry flounder, Platichthys stellatus. Aquacult 433, 105-114.
- Maita, M. (2007) Fish Health Assessment In Dietary Supplements for the Health and Quality of Cultured Fish (Nakagawa H., S.M., Gatlin D. M. ed.), Cab International pp: 10-34.
- Mallekh, R., Lagardère, J.P., Bégout Anras, M.L., Lafaye, J.Y. (1998) Variability in appetite of turbot, Scophthalmus maximus under intensive rearing conditions: the role of environmental factors. Aquaculture 165, 123-138.
- Mas-Muñoz, J., Komen, H., Schneider, O., Visch, S.W., Schrama, J.W. (2011) Feeding Behaviour, Swimming Activity and Boldness Explain Variation in Feed Intake and Growth of Sole (Solea solea) Reared in Captivity. Plos one 6, e21393.
- Meena, D.K., Das, P., Kumar, S., Mandal, S.C., Prusty, A.K., et al. (2013) Beta-glucan: an ideal immunostimulant in aquaculture (a review). Fish Physiol and Biochem 39, 431-457.
- Misra, C.K., Das, B.K., Mukherjee, S.C., Pattnaik, P. (2006) Effect of long term administration of dietary β-glucan on immunity, growth and survival of Labeo rohita fingerlings. Aquaculture 255, 82-94.
- Nakagawa, H., Sato, M., Gatlin, D.M. (2007) Dietary Supplements for the Health and Quality of Cultured Fish.
- Okorie, O.E., Bae, J.Y., Kim, K.W., Son, M.H., Kim, J.W., et al. (2013) Optimum feeding rates in juvenile olive flounder, Paralichthys olivaceus, at the optimum rearing temperature. Aquaculture Nutrition 19, 267-277.
- Peng, M., Xu, W., Ai, Q., Mai, K., Liufu, Z., et al. (2013) Effects of nucleotide supplementation on growth, immune responses and intestinal morphology in juvenile turbot fed diets with graded levels of soybean meal (Scophthalmus maximus L.). Aquaculture 392, 51-58.
- Peng, M., Xu, W., Mai, K., Zhou, H., Zhang, Y., et al. (2014) Growth performance, lipid deposition and hepatic lipid metabolism related gene expression in juvenile turbot (Scophthalmus maximus L.) fed diets with various fish oil substitution levels by soybean oil. Aquaculture 433, 442-449.
- Pham, M.A., Byun, H.G., Kim, K.D., Lee, S.M. (2014) Effects of dietary carotenoid source and level on growth, skin pigmentation, antioxidant activity and chemical composition of juvenile olive flounder Paralichthys olivaceus. Aquaculture 431, 65-72.
- Rana, K.J., Siriwardena, S., Hasan, M.R. (2009) Impact of rising feed ingredient prices on aquafeeds and aquaculture production p: 63.
- RingØ, E., Olsen, R.E., Gifstad, T.Ø., Dalmo, R.A., Amlund, H., et al. (2010) Prebiotics in aquaculture: a review. Aquaculture Nutrition 16, 117-136.
- Sakai, M. (1999) Current research status of fish immunostimulants. Aquaculture 172, 63-92.
- Song, Z., Li, H., Wang, J., Li, P., Sun, Y., et al. (2014) Effects of fishmeal replacement with soy protein hydrolysates on growth performance, blood biochemistry, gastrointestinal digestion and muscle composition of juvenile starry flounder (Platichthys stellatus). Aquaculture 426, 96-104.
- Staykov, Y., Spring, P., Denev, S., Sweetman, J. (2007) Effect of a mannan oligosaccharide on the growth performance and immune status of rainbow trout (Oncorhynchus mykiss). Aquaculture International 15, 153-161.
- Torrecillas, S., Makol, A., Caballero, M.J., Montero, D., Dhanasiri, A.K.S. (2012) Effects on mortality and stress response in European sea bass, Dicentrarchus labrax (L.), fed mannan oligosaccharides (MOS) after Vibrio anguillarum exposure. J Fish Diseases 35, 591-602.
- Bussel, C.G.J.V, Mahlmann, L., Kroeckel, S., Schroeder, J.P., Schulz, C. (2013) The effect of high ortho-phosphate water levels on growth, feed intake, nutrient utilization and health status of juvenile turbot (Psetta maxima) reared in intensive recirculating aquaculture systems (RAS). Aquacultural Engineering 57, 63-70.
- Bussel, C.G.J.V, Schroeder, J.P., Wuertz, S., Schulz, C. (2012) The chronic effect of nitrate on production performance and health status of juvenile turbot (Psetta maxima). Aquaculture 326, 163-167.
- Wang, J., Li, B., Ma, J., Wang, S., Huang, B., Sun, Y., Zhang, L. (2015) Optimum dietary protein to lipid ratio for Starry flounder (Platichthys stellatus). Aquaculture Research.
- Wang, J., Zhang, D., Sun, Y., Wang, S., Li, P., Gatlin, D.M., Zhang, L. (2016) Effect of a dairy-yeast prebiotic (GroBiotic®-A) on growth performance, body composition, antioxidant capacity and immune functions of juvenile starry flounder (Platichthys stellatus). Aquaculture Research 47, 398-408.
- Wang, Y.B., Li, J.-R., Lin, J. (2008) Probiotics in aquaculture: Challenges and outlook. Aquaculture 281, 1-4.
- Yoo, G., Lee, S., Kim, Y.C., Okorie, O.E., Park, G.J., et al. (2007) Effects of Dietary β-1,3 Glucan and Feed Stimulants in Juvenile Olive Flounder, Paralichthys olivaceus. J World Aquacult Society 38, 138-145.
- Zekovi, D.B., Kwiatkowski, S., Vrvi, M.M., Jakovljevi, D., Moran, C.A. (2005) Natural and Modified (1→3)-β-D-Glucans in Health Promotion and Disease Alleviation. Critical Reviews in Biotechnology 25, 205-230.


