Keywords
Sterlet, Feed conversion, Meat yield, Feed content
Introduction
The sterlet, Acipenser ruthenus, is a freshwater species. It originally inhabits the rivers of Eurasia, being widely distributed in rivers flowing into the Caspian, Black, Baltic, White, Barents and Kara Seas and the Sea of Azov. They reach to sexual maturity at 3-5 years in males and 5-8 years in females in nature condition (Hochleithner and Gessner, 1999).
The optimal temperature of sterlet is 18-22°C in 200-250 g and its growth rate decreases rapidly below 14°C (Ponomareva et al., 2002). It is a benthos feeder and its diet changes according to season and river inhabitants, but generally includes insect larvae, small molluscs and invertebrates (Solokov and Vasiliev, 1989).
The captive breeding of sturgeons was commenced in 1869 by a Russian academician Ovsyannikov who successfully fertilized sterlet eggs using milt of the Russian sturgeon (A. gueldenstaedtii) and stellate sturgeon (A. stellatus) (Ovsyannikov, 1870, 1872, 1873).
The most commonly used species is the Siberian sturgeon (Acipenser baerii) which is presently reared in 22 countries reaching a total production of about 8800 tonnes per year, followed by the Russian sturgeon cultured in about 16 countries, while the sterlet is presently cultured in 15 countries (Bronzi et al., 2011).
There is a few studies dealing with the culture of sturgeon (Williot et al. 2005; Gencharov, et al., 2009; Ronyai, 2009) and new studies continue to provide experience about sterlet reproduction reproduction. As a freshwater species sterlet is one of the species identified as a leading candidate for sturgeon culture, because it reaches commercial weight (1-2 kg) and sexual maturity (3-4 years) fastly. Under recirculation system conditions it can be propagated at the age of 2-3 years with an inter-spawning period of 1 year (Ronyai, 2006a; Ponomareva and Matishov, 2006).
The studies on the nutritional requirements of the sturgeons are mostly related to white sturgeon (Acipenser transmontanus) and Siberian sturgeon (Moore et al., 1988; Hung et al., 1989; Kaushik et al., 1989; Xu et al., 1993; Deng et al., 1998; Stuart and Hung, 2003; ?ener et al., 2005). The previous literature for some cultured sturgeon species was revived and summarized by García- Gallego et al. (2009). In published literature plenty of studies exist on rearing of sturgeon species. Some of those studies focus on growth rates of white sturgeon (Stuart and Hung, 2003), Siberian sturgeon (Ronyai et al., 1989a; Rad et al., 2003; Adámek et al., 2007), sterlet (Ronyai et al., 1989b; Jecu, et al., 2008; Feledi et al., 2011; Prokes et al., 2011) and Russian sturgeon (Çelikkale et al., 2002; Memi? et al., 2009), some of them are about feeding rates or frequency of feeding of white sturgeon (Hung and Lutes, 1987; Hung et al., 1993), Siberian sturgeon (Ronyai et al., 1989b; Ronyai et al., 1999; Köksal et al., 2002), Russian sturgeon (Memi? et al., 2009), great sturgeon, Huso huso (Mohseni et al., 2006), juvenile Atlantic sturgeon (Acipenser oxyrinchus) and shortnose sturgeon, A. brevirostrum (Giberson and Litvak, 2003). There are also a number of publications dealing with a stock density for white sturgeon (Gershanovich and Taufik, 1992). However, studies on the feeding rate of sterlet and its effects on growth perfor-mance and feed utilization are scarce (Ronyai et al., 2006; Feledi et al., 2011).
In intensive fish farming operational cost mostly depends on the feed because it accounts for 60-80 percent of total production costs (Hasan et al., 2007). Therefore feeds will continue to dominate fish farming needs. On the other hand, growth performance and food utilization are ma-jor variables in fish culture and mostly related to water temperature. The aim of any fish farming is using low cost feeds and obtaining better growth performance.
There were also several studies about meat yield of different fish species (Macias 2004; Samsun et al., 2005; Duman and Dartay, 2007; ?ebnem et al., 2011) and effects of cultivation practices on it (Cibert et al., 1999).
The objective of the present study is to inves-tigate the effect of different feeding rates with a low fat diet containing appropriate protein on growth performance, food utilization and meat yield of sterlet reared in a recirculating aquacul-ture system.
Materials and Methods
The fish used in the experiment hatched in April 2011 and raised in the Research Institute for Fisheries, Aquaculture and Irrigation (HAKI) in Szarvas, Hungary. The experiment was con-ducted in January-Ferbuary 2012. Prior to meas-urement the fish was anaesthetized with clove oil (Akbulut et al., 2011). 120 fish with an average weight of 187 g were individually weighed to nearest 0.1 g and randomly placed into six tanks (1.1x1.1 m) in 2 subgroups. Water depths in the tanks with a middle outflow were set up to 25 cm with a 10 L/minute flow rate.
Six groups composing 20 sterlets each were fed with a commercial (3-3.5 mm) sunken pellet feed (Halpad Ltd. Szarvas, Hungary) with 45% protein and 6.5 % fat content. Three feeding rates were applied: 1, 2 and 3% of total body weight per day (BW/day), and the daily rations were cal-culated with expecting 0.6 SGR (%). The amounts of feed were correlated by weight meas-urements every 20 days. The feeding commenced at 08:00 every day at belt feeders set for 12 hours and lasted for 60 days. During the experiment water quality parameters were also checked, in-cluding daily temperature records with a mini-mum of 16.6°C and a maximum of 19.9°C (aver-age temperature was 18.7±0.97°C). The other measurements varied as follows: oxygen concen-tration 7.9-9.4 mg/l and saturation 81-99%.
Percentage relative change in weight was also used: RW change (%)=100 x (Wf–Wi)/Wi where t is the number of experiment days. Wi and Wf are the initial and final weights (g), F is the cumula-tive food consumption of fish in each tank.
The coefficient of variation (CV) was used to examine an inter-individual weight variation among the fish in each group: CV (%)=100xS.D./mean weight of the fish in each tank. Percentage relative was calculated as: CV change (%)=100*(CVf–CVi)/CVi.
At the end of experiment five fish from each tank were randomly sampled for indices estima-tions. Body weights, livers and viscera were indi-vidually weighed for calculation of hepatosomat-ic index (HIS), viscerasomatic index (VSI) and carcass efficiency (CE). Viscera included a liver and a gastrointestinal tract from esophagus to anus. Carcass weight (CW) was determined by subtracting viscera, head and fins from body weight.
The following variables were calculated: hepatosomatic index HSI= [liver weight(g)/body weight (g)] x 100, viscera somatic index VSI=[viscera weight (g)/body weight (g)] x 100) and carcass efficiency CE= [Carcass weight (g)/body weight (g)]x100.
Statistical tests were carried out using the SPSS software package for Windows. All data were subjected to one-way analysis of variance (ANOVA) and differences between means com-pared by the Tukey test at a 95% confidence in-terval (P<0.05).
Results and Discussion
Feeding rate affected the growth of the fish. Feeding rate for maximum growth was 2% of BW/day. Mean weight gains were 75.4±4.95 g, 98.7±6.40 g, and 94.1±5.76 g for fish in the 1, 2 and 3% of BW/day groups, respectively. The mean weight increase in the 1 % group was sig-nificantly lower than the other groups (P<0.05).
Growth trend of fish during the course of the ex-periment is shown in Figure 1. The specific growth rates (SGR%) were significantly higher (P<0.05) in the 2 and 3% feeding rate groups than in 1% group (Table 1). Statistically signifi-cant differences were found in FCR between the 1, 2 and 3% of BW/day groups (P>0.05). The feeding rate did not influence neither the hepato-somatic index (HIS), nor viscerasomatic index (VSI) and carcass efficiency (CE) (P>0.05). The final meat yield parameters of sterlet are repre-sented in Table 2.

Figure 1. Average weight of juvenile sterlets fed with 1, 2 and 3 percent of BW/day.
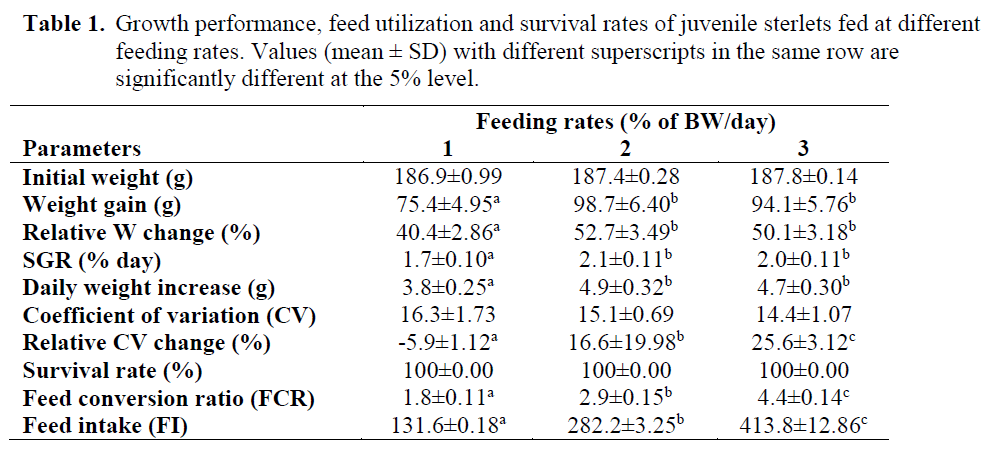
Table 1. Growth performance, feed utilization and survival rates of juvenile sterlets fed at different feeding rates. Values (mean ± SD) with different superscripts in the same row are significantly different at the 5% level.
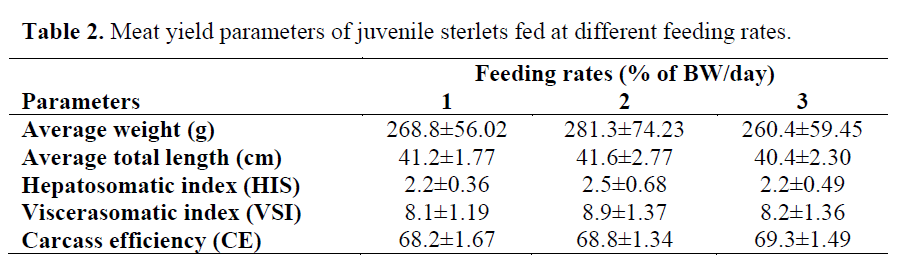
Table 2. Meat yield parameters of juvenile sterlets fed at different feeding rates.
In sturgeon culture, feeding rates vary accord-ing to the purpose and stage of rearing systems (Ronyai, 2006a; Chebanov and Galich, 2010; Prokes et al., 2011). Besides, different diet con-tent is recommended for sturgeon feeding de-pending on the fish size and species (Hung and Deng, 2002; Chebanov and Galich, 2010).
The results obtained in the present study were similar to those in the study by ?ener et al. (2006) where a feeding rate of 2% body weight was used at 16°C for Russian sturgeon with average weight of 144 g and studies of Sion et al. (2011) who used a 41 g sterlet fed with 30-46% crude protein at the 1.5% BW/day feeding rate at 19.1-24.6°C temperature.
Our SGR results in all the experimental groups were higher than results of lower tempera-ture studies of Memi? et al. (2006) who found SGR as 0.16 for Russian sturgeon fed with a carp diet containing 35% protein and 10% crude fat, and Akbulut et al. (2010) where SGR was found as 0.36 for 0.5-2.2 kg juvenile beluga fed with raw fish at 13°C.
Filipiak et al. (1999) reported that SGR and FCR were 3.03 and 1.57 for 25 g sterlet reared in cages at 12.2-23.0 °C with feeding a diet contain-ing 41.3% protein and 23.3% fat. Feledi et al. (2011) found SGR and FCR as 5.39 and 3.23 in 77 g sterlet reared in the tanks at 24.0-25.6 °C feeding with a diet containing 45% protein and 6.5% fat at feeding rate of 10% BW. According to Prokes et al. (2011), during the subsequent part of juvenile period of sterlet, from the 6th day after hatching and up to the age 5+ the SGR values subsequently diminish from 7.00 to 0.05 under farm conditions, and SGR summers in 0.88, 0.73 and 0.59, FCR in 1.35, 1.39 and 1.42, with daily feeding rates 1.6, 1.7 and 2.1 % for 99.5 and 71.2 g juvenile sterlets, respectively.
When comparing growth and feed utilization of sterlet with other sturgeon species, it can be seen that growth rate of sterlet was a little bit slower but feed utilization was similar to those of the other sturgeons. In the case of meat yield of sterlet there are no published data, but there are some numbers for Russian sturgeon, for example, Sener et al. (2005 and 2006) who reported that dietary lipid effected on values of HIS and VSI in Russian sturgeon. Akbulut et al. (2013) calculat-ed HIS, VSI and CA as 3.28, 8.18 and 65.14 for 400 g Russian sturgeon and 2.97, 9.03 and 65.40 for stellate sturgeon.
Conclusion
Consequently determining the optimal feeding rates for sterlet is important to the success of its culture. Meat yield may also be helpful to deter-mine a daily feed amount of food in sturgeon cul-ture.
Acknowledgements
The financial support was provided by the Republic of Turkey, Ministry of Food, Agricul-ture and Livestock for this study which was con-ducted at the Research Institute for Fisheries, Aquaculture and Irrigation, Szarvas, Hungary. Many thanks to all our colleagues who helped and supported us during the study.
454
References
- Adámek, Z., Prokeš, M., Baruš, V., Sukop, I., (2007). Diet and Growth of 1+ Siberian Sturgeon, Acipenserbaerii in Alternative Pond Culture, Turkish Journal of Fisheries and Aquatic Sciences, 7: 153-160
- nAkbulut, B., Çakmak, E., Kurtoglu, I.Z. ve Ak-sungur, N., (2010). Karadeniz’denYakalan-anveTanklardaMezgi tile BeslenenGenç Mersin Morinalarinin (HusoHuso) BüyümePerformansi, Journal of FisheriesScienc-es.com, 4(2): 184-189.doi: 10.3153/jfscom.2010019
- nAkbulut, B., Çakmak, E., Aksungur, N., Çavdar, Y., (2011). Effect of Exposure Duration on Time to Recovery from Anaesthesia of Clove Oil in Juvenile of Russian Sturgeon, Turkish Journal of Fisheries and Aquatic Sciences, 11: 463-467. doi: 10.4194/1303-2712-v11_3_17.doi: 10.4194/1303-2712-v11_3_17
- nAkbulut, B., Aksungur, N., Çakmak, E., Kutlu, S., Yesilsu, A. F., Alp, E., (2013). Effect of the Feeding Frequency with Restricted Feed-ing Rate on Growth, Feed Conversion and Body Composition of Juvenile Russian and Stellate Sturgeon in Recirculating System, Journal of Applied Ichthyology, (In pro-gress). 10.1111/j.1439-0426.2011.01757.x
- nBronzi, P., Rosenthal, R., Gessner, J., (2011). Global sturgeon aquaculture production: an overview, Journal of Applied Ichthyology, 27: 169-175. doi:10.1111/j.1439-0426.2011.01757.x
- nChebanov, M.S., Galich, E.V., (2010). Sturgeon Hatchery Manual, FAO Fisheries and Aqua-culture Technical Paper 558, Ankara, pp. 306
- nDuman, M., Dartay, M., 2007. Changes in meat yield and chemical composition of mirror carp (Cyprinuscarpio L.,1758) fillets after hot-smoked, DoguAnadoluBölgesiArastirmalari, 5(3): 186-190
- nCibert, C., Fermon, Y., Vallod, D., Meunier, F.J., (1999). Morphological screening of carp, C. carpio: relationship between morphology and fillet yield, Aquatic Living Resources, 12(1): 1-10.doi: 10.1016/S0990-7440(99)80009-6
- nÇelikkale, M.S., Timur, M., Memis, D., Ercan, E., (2002). Influence of Acclimation to the Cold Water on Growth Rate of Russian Sturgeon Juveniles (Acipensergueldenstaed-tii, Brandt&Ratzenburg, 1833), Turkish Journal of Fisheries and Aquatic Sciences, 2(2): 137-140
- nDeng, D.F., Hung, S.S.O., Conklin, D.E., (1998). White sturgeon (Acipensertransmontanus) require both n-3 and n-6 fatty acids, Aqua-culture, 161: 333-335.doi: 10.1016/S0044-8486(97)00280-9
- nFeledi, T., Lengyel, S., Ronyai, A., (2011). Pre-liminary Results of Intraspecific Sterlet Hy-brid (Siberian starlet x Sterlet) Rearinig Un-der Intensive Conditions, Aquaclture in Cen-tral and Eastern Europe: present and Fu-ture, The II. Assembly NACEE. 274-76
- nFilipiak, J., Czerniejewski, P., Sadowski, J., Trzebiatowski, R., (1999). Comparison of The Effects of Cage–Rearing of Sterlet (Acipenserruthenus) and Russian × Siberian Sturgeon (Acipensergueldenstadetdti × A.baeri) Hybrid Fry in Cooling Water, Electronic Journal of Polish Agricultural Universities, 2(2): 03
- nGarcía-Gallego, M., de la Higuera, M., Hidalgo, M.C., Furné, M., Morales, A.E., Sanz, A., (2009). On Nutrition and Feeding Studies as the Basis for the Culture of Different Stur-geon Species. In: Carmona, R., Domezain, A., García-Gallego, M., Hernando, J.A.,Rodríguez, F. and Ruiz-Rejón, M. (Eds), Bi-ology, Conservation and Sustainable Devel-opment of Sturgeons, Springer Science Business Media B.V. pp. 215-234
- nGershanovich, A.D., Taufik, L.R., (1992). Feed-ing dynamics of sturgeon fingerlings Acipenseridae. Depending on food concen-tration and stocking density, Journal of Fish Biology, 41: 425-434.doi: 10.1111/j.1095-8649.1992.tb02671.x
- nGiberson, A.V., Litvak, M.K., (2003). Effect of Feeding Frequency on Growth, Food Con-version Efficiency, and Meal Size of Juve-nile Atlantic Sturgeon and ShortnoseStur-geon, North American Journal of Aquacul-ture, 65: 99-105.doi: 10.1577/1548-8454(2003)65<99:EOFFOG>2.0.CO;2
- nHasan, M.R., Hecht, T., De Silva, S.S., Tacon, A.G.J., (2007). Study and analysis of feeds and fertilizers for sustainable aquaculture development. Fisheries Technical Paper, 497, Rome, pp. 531
- nHochleithner, M., Gessner, J., (1999). The Stur-geon and Paddlefishes (Acipenseriformes) of the World: Biology and Aquaculture, AquaTech Publications. pp. 165
- nHung, S.S.O., Lutes, P.B., (1987). Optimum feeding rate of hatchery-produced juvenile White sturgeon (Acipensertransmontanus): at 20 degree C, Aquaculture, 65(3-4): 307-317.doi: 10.1016/0044-8486(87)90243-2
- nHung, S.S.O., Fynn-Aikins, K., Lutes, P.B., Xu, R., (1989). Ability of juvenile White stur-geon (Acipensertransmontanus) to utilize different carbohydrate sources, Journal of Nutrition, 119: 727-733
- nHung, S.S.O., Lutes, P.B., Shqueir, A.A., Conte, F.S., (1993). Effects of feeding rate and wa-ter temperature on growth of juvenile white sturgeon (Acipencertransmontanus), Aqua-culture, 115: 297-303.doi: 10.1016/0044-8486(93)90144-N
- nHung, S.O., Deng, D.F., (2002). Sturgeon, Acipenser spp. In: Webster, C.D., Lim, C.E. (Eds), Nutrient Requirements and Feeding of Finfish for Aquaculture. CABI Publishing New York, pp. 34-358
- nJecu, E., Patriche, N., Talpes, M., Paltenea, E., Trofimov, A., Esanu, V., (2008). Technolog-ical Aspects Regarding Rearing of The Acipenserruthenus Species, Albino Variety to Second Summer Old, IN Brates Sturgeons Station, LucraristiinNificeZootehniesi Bio-tehnologii, 41 (2): 69-74
- nKaushik, S.J., Luquet, P., Blanc, D., Paba, A., (1989) Studies on the nutrition of Siberian sturgeon, (Acipenserbaeri). In. Utilization of digestible carbohydrates by sturgeon, Aq-uaculture, 76: 97-107.doi: 10.1016/0044-8486(89)90254-8
- nKöksal, G., Rad, F., Kindir, F.M., (2002). Growth Performance and Feed Conversion Efficiency of Siberian Sturgeon Juveniles (Acipenserbaeri) Reared in Concrete Race-ways, Turkish Journal of Veterinary and An-imal Sciences, 24: 435-442
- nMacias, J.A.G., González, F.A.N., Pineda, O.C., Rodríguez, R.H.A., Hernández, M.R.E., (2004). Carcass and meat quality of rainbow trout, Oncorhynchusmykiss Richardson, produced in the northwest of Chihuahua State, Hidrobiologica, 14(1): 19-26
- nMemis, D., Çelikkale, M., Ercan, E., (2006). Ef-fects of different diets on growth perfor-mance and body composition of Russian sturgeon (Acipensergueldenstaedtii, Brandt&Ratzenburg, 1833), Journal of Ap-plied Ichthyology, 22: 287-290.doi: 10.1111/j.1439-0426.2007.00970.x
- nMemis, D., Ercan, E., Çelikkale, M.S., Timur, M., Zarkua, Z., (2009). Growth and Survival Rate of Russian Sturgeon (Acipenserguel-denstaedtii) Larvae from Fertilized Eggs to Artificial Feeding, Turkish Journal of Fish-eries and Aquatic Sciences, 9: 47-52
- nMohseni, M., Pourkazemi, M., Bahmani, M., Falahatkar, B., Pourali, H.R., Salehpour, M., (2006). Effects of feeding rate and frequency on growth performance of yearling great sturgeon, Husohuso, Journal of Applied Ichthyology, 22: 278-283.doi: 10.1111/j.1439-0426.2007.00968.x
- nMoore, B.J., Hung, S.S.O., Medrano, J.F., (1988). Protein requirement of hatchery-produced juvenile white sturgeon (Acipensertransmontanus), Aquaculture, 71(3): 235-245.Ovsyannikov, F.V., (1870). On the artificial breeding of sterlet. Trudy 2-go S'ezda Russ-kikhEstestvoispytateleipoOtdelamZo-ologii, Anatomii, Fiziologii
- nOvsyannikov, F.V., (1872). On the sterletartifi-cial breeding experiments. Trudy Impera-torskogoVol'no-EkonomicheskogoObshchestva2, No. 1
- nOvsyannikov, F.V., (1873). The first experiment on the artificial breeding of sterlet in the Sankt-Petersburg region. Trudy Sankt-PetersburgskogoObshchestvaEstestvoispytatelei 4, No. 2
- nPonomareva, S., Gamygin, E., Nikonorov, S., (2002). Technologies of aquaculture objects rearing and feeding in Southern Russia. As-trakhan, Nova Plus. pp. 264 (in Russian)
- nPonomareva, E., Matishov, G., (2006). Cultiva-tion the Don sterlet in industrial conditions, International Conference Aqua 2006, Flor-ence, Italy
- nProkes, M., Baruš, V., Mareš, J., Penáz, M., Ba-ránek, V., (2011).Growth of starlet Acipenserruthenus under experimental and farm conditions of the Czech Republic, with remarks on other sturgeons. Actauniv. agric. et silvic. Mendel. Brun., 2011, LIX, 6: pp. 281-290
- nRad, F., Köksal, G., Kindir, M., (2003). Growth Performance and Food Conversion Ratio of Siberian Sturgeon (Acipenserbaerii Brandt) at Different Daily Feeding Rates, Turkish Journal of Veterinary and Animal Sciences, 27: 1085-1090
- nRonyai, A., Ruttkay, A., Varadi, L., Peteri, A., (1989a). Growth of Siberian sturgeon (Acipenserbaerii L.) and that of its hybrids with the sterlet (Acipenserruthenus in recy-cling systems). In: Williot, P. (Ed.), Acipenser, CEMAGREF, 423-428
- nRonyai, A., Ruttkay, A., Varadi, L., Peteri, A., (1989b). Growth comparative trial of finger-lings of sterlet (Acipenserruthenus L.) and its hybrids with male Siberian sturgeon (Acipenserbaerii B). In: Williot, P. (Ed.), Acipenser, CEMAGREF, 417-422
- nRonyai, A., (1999). Effect of ration size and wa-ter temperature on growth of juvenile Sibe-rian sturgeon (Acipenserbaeri Brandt), Journal of Applied Ichthyology, 15: 344-345. doi: 10.1111/j.1439-0426.1999.tb00373.x
- nRonyai, A., (2006a). Artificial propagation and larval rearing of sterlet (Acipenserruthenus L.) International Workshop on Sturgeon Conservation and Breeding, Szarvas, Hun-gary
- nRonyai, A., (2006b). A growth model for the practical use in sturgeon feeding, Interna-tional Conference Aqua 2006, Florence, Ita-ly
- nRonyai, A., (2009). The effect of different syn-theticgonadotrop-releasing hormone ana-logues and their combinations with an anti-dopaminergic compound on the reproduc-tion performance of sterlet (Acipenserruthenus L.), Aquaculture Research, 40: 315-322. doi: 10.1111/j.1365-2109.2008.02094.x
- nSamsun, N., Samsun, O., Kalayci, F., (2005). Seasonal Variations of Meat Yield and Pro-tein& Oil Rates of Turbot (Scophthalmusmaeoticus Pallas, 1811) Caught in Sinop Region (Black Sea), Science and Enginee-ring Journal of Firat University, 17 (4): 629-635
- nSion, C., Calin, P.G., Oprea, L., Nica, A., Bacanu, M.G., (2011). The influence of pel-lets quality on the growth of sterlet, in recir-culating aquaculture system, AACL Bioflux, 4(2): 129-130
- nSolokov, L.I., Vasiliev, V.P., (1989). AcipenserruthenusLinneaus. In: Holcik,J. (Ed), The Freshwater Fishes of Europe, Vol.I-II AULA-Verlag, Weisbaden, 227-262
- nSebnem, A. S., Basçinar, N., Kocabas, M., Tufan, B., Köse, S., Okumus, I., (2011). Evaluation of Meat Yield, Proximate Composition and Fatty Acid Profile of Cultured Brook Trout (SalvelinusfontinalisMitchill, 1814) and Black Sea Trout (Salmotruttalabrax Pallas, 1811) in Comparison with their Hybrid, Turkish Journal of Fisheries and Aquatic Sciences, 11: 261-271.doi: 10.4194/trjfas.2011.0211
- nSener, E., Yildiz, M., Savas, E., (2005). Effects of Dietary Lipids on Growth and Fatty Acid Composition in Russian Sturgeon (Acipensergueldenstaedtii) Juveniles, Turk-ish Journal of Veterinary and Animal Sci-ences, 29: 1101-1107
- nSener, E., Yildiz, M., Savas, E., (2006). Effect of Vegetable Protein and Oil Supplementation on Growth Performance and Body Composi-tion of Russian Sturgeon Juveniles (Acipensergueldenstaedtii Brandt, 1833) at Low Temperatures. Turkish Journal of Fish-eries and Aquatic Sciences, 6: 23-27
- nStuart, J.S., Hung, S.S.O., (2003). Growth of juvenile white sturgeon (Acipensertransmon-tanus) fed different proteins, Aquaculture, 78(3-4): 303-316
- nWilliot, P., Brun, R., Rouault, T., Pelard, M., Mercier, D., Ludwig, A., (2005) Artificial spawning in cultured sterlet sturgeon, Acipenserruthenus L., with special empha-sis on hermaphrodites, Aquaculture, 246: 263-273.doi:10.1016/j.aquaculture.2005.02.048
- nXu, R., Hung, S.S.O., German, J.B., (1993). White sturgeon tissue fatty acid composi-tions are affected by dietary lipids, Journal of Nutrition, 123(10), 1685-1692.


