Keywords
Nile tilapia; Sexual steroid hormone; Estrogen and androgen receptor genes
Introduction
Tilapia are increasingly important to global food security. China occupies approximately 30% of global tilapia products and is a lead producer (Yue, Zhao & Tang 2017). Fish generally exhibit a variety of sexual dimorphism, which is regulated by sex hormones that are involved in several physiological processes, including reproduction, growth, and development (Phumyu, Boonanuntanasarn & Jangprai 2012). Sex steroid hormones (e.g., androgens and estrogens) mediate their action on gene expression via their cognate receptors. The classical mechanism of steroid hormone action involves ligand binding, receptor dimerization, nuclear translocation, binding to hormone response elements, recruitment of coregulators, and chromatin remodeling, all of which lead the regulation of gene transcription (Lee & Chang 2003; Smith & Walker 2014).
The induction of 17β-estradiol (E2) functions primarily through two cognate receptors, estrogen receptor (ER) α and β (White, Lees, Needham, Ham & Parker 1987; Kuiper, Enmark, Peltohuikko, Nilsson & Gustafasson 1996). After binding to estrogen, the ER interacts with a specific response element on DNA, thereby promoting the expression of target genes or proteins (Shupnik & Rosenzweig 1991).
Androgens mediate a wide range of male developmental processes, and are particularly important in male sexual differentiation (Heinlein & Chang 2002). Testosterone (T) exerts its function predominately through androgen receptor (AR)- mediated pathways (Lee et al., 2003).
The AR is a steroid receptor for testosterone (T), which belongs to the nuclear receptor super-family and exerts its effects as a transcription factor which regulates the activity of target genes (Heinlein et al., 2002); however, the mechanism by which these genes are regulated by estrogen and androgens in female and male fish remain unclear. Western blotting and immunohistochemical staining are common techniques for determining protein expression in specific tissues. In the present study, we evaluated the effect of ER and AR on gene expression, protein expression, and positive immunohistochemical reaction in the liver, intestine, gonad, and kidney tissues of male and female tilapia. Identifying how sex steroids affect ER and AR mechanisms in female and male fish improve the capacity to understand sexual dimorphism and potential interaction between growth and reproduction.
Materials and Methods
Fish
Male and female Nile tilapia (100 g ± 5 g) were reared in freshwater semi-recirculating tanks at Shanghai Ocean University, China. Body weight was recorded to the nearest 0.1 g. Prior to the experiment, fish were fed a known amount of feed (special feed for tilapia “FFEF3”, Ningbo Tianbang Company, China) twice daily at 10:00 and 17:00.
The fish were randomly divided into groups of females and males. Each group consisted of 24 fish, which were assigned to one of three treatments: 1) E2; 2) T; 3) control. Sex steroid hormones (SIGMA-ALDRICH, China) were resuspended (100 μg/μL) in 100% ethanol and diluted to 25 μg/μL in vegetable oil. The control treatment received a ratio of ethanol:vegetable oil equivalent to that of the steroid suspensions. The final steroid dosages were 50 μg/g body weight (Yue et al., 2017). At 18:00, experimental fish (excluding control fish) were anesthetized with sodium bicarbonate-buffered MS-222 (50 mg/L, weighed to calculate the injection volume), injected with sex steroid hormones, and placed immediately back in the tanks. Liver, gonad, kidney, and intestine samples were collected 24 h post-injection, snap frozen in liquid nitrogen, and stored at -80°C.
RNA extraction and DNase treatment
Total RNA was extracted from the indicated tissues (50 mg-100 mg) using TRIzol reagent (Invitrogen, Carlsbad, CA) according to the standard protocol. RNA quality and quantity were determined at 260 nm and 280 nm, respectively (Colibri, Berthold Technologies GmbH & Co. KG Germany). Prior to reverse transcription, a portion of each RNA sample was treated with 10 U RNase-free DNase I (20 μg-50 μg total RNA per 50 μL reaction mix volume; TaKaRa Bio, Otsu, Shiga, Japan) according to the manufacturer's standard protocol to remove any genomic DNA.
Reverse transcription-polymerase chain reaction (RT-PCR)
For cDNA synthesis, 10 μg of total RNA was reversetranscribed using a high-capacity cDNA reverse transcription kit (TaKaRa, Japan). Gene expression was analyzed by real-time RT-PCR (Bio-Rad iQ5 Real-time PCR System, Bio-Rad, USA), using TransStart™ Green qPCR SuperMix (TaKaRa, Japan). Each sample was analyzed in duplicate. Gene-specific primers (Table 1) were designed using the Primer-BLAST tool from NCBI. The expression level of the GH/IGF (growth hormone insulin-like growth factor) axis and MRF (muscle regulatory factors) genes were expressed relative to those of β-actin.
| Gene symbol |
Gene name |
Primer sequence |
GeneBank accession No. |
| Estrogen receptor |
| ER-β |
estrogen receptor-β |
5’-CATCCCTCCTACCCTGTCTTA |
AM284391 |
| 5’-TCCAGGTATTTGAAGGTCCGC |
| Androgen receptor |
| AR-α1 |
androgen receptor-α1 |
5’-GGAGCTGCAAGTTTCTTCAAAGAGC |
NM_001279613 |
| 5’-CCGGCACTTGGCCCACTTGACAC |
| β-actin |
Reference gene |
5’-CAGCAGATGTGGATCAGCAAGC |
|
| 5’- TGAAGTTGTTGGGCGTTTGG |
Table 1: Genes of interest and primers used in real-time qPCR. Primer sequences were designed according to GenBank.
For PCR, the reaction volume (15 μL) contained 0.5 μL of each primer (10 μM), 1 μL of 10-1 diluted cDNA, 3 μL PCR-grade water, and 10 μL of 2× TransStart™ Green qPCR SuperMix. Samples were analyzed in triplicate using the Bio-Rad iQ5 Real-time PCR System (Bio-Rad, CA, USA). The PCR conditions consisted of one cycle at 95°C for 5 min, followed by 45 cycles at 94°C for 15 s, 65°C for 15 s, and 72°C for 15 s (Livak & Schmittgen 2001).
RT-PCR data were analyzed using CFX Manager TM software (Version 1.0). The level of gene expression was calculated using the 2−ΔΔCt method, in which ΔCt represents differences in the Ct values between the gene of interest and β-actin. The ΔΔCt was the ΔCt for each sample subtracted from that of the calibrator.
Western blotting
According to the method of Kumar, Yadav, and Pakrasi (2016), Western blots were performed with a slight modification and the help of the blue color impression of pontamine dye (Sigma, USA). The different tissues were homogenized in lysis buffer containing 150 Mm NaCl, 50 Mm Tris (pH 8), 1% NP-40, 0.1% SDS, 0.5% sodium deoxycholate, and fresh protease inhibitor (1 μg/mL aprotinin, 1 mM EDTA, 0.5 μg/mL leupeptin, 100 μg/mL PMSF, and 1 mM sodium orthovanadate). Tissue homogenates were kept on a rocker at 4°C for 30 min. The homogenates were centrifuged at 10,000 × g at 4°C for 10 min and the supernatants were collected and stored at -80°C. The protein concentration was determined using a Quick Start Bradford Protein Assay (Bio-Rad, USA). A 35 μg sample of each protein was boiled in 2 × Laemmli sample buffer (4% SDS, 20% glycerol, 125 mM Tris [pH 6.8] 0.02% bromophenol blue, and 10% β-mercaptoethanol) for 5 min. The proteins were subjected to 10% SDS-PAGE and transferred to a PVDF membrane (Millipore, USA). The membranes were blocked in 5% non-fat milk in TBS-T (10 mM Tris-HCl [pH 8] and 150 mM NaCl and 0.05% tween 20) for 2 h at room temperature. The membranes were incubated with primary antibodies for ER (ab288, 1:800), AR (Abcam, 1:500), and GAPDH (Weiao, 1:2000) overnight at 4°C (abcam, USA). Following incubation, the membranes were washed twice (10 min each) with TBS-T and subjected to an incubation with a secondary antibody (antirabbit Jackson 1:2000) incubation in 5% milk for 2 h at room temperature. Incubation in 5% milk for 2 h at room temperature.
Immunohistochemistry
The samples obtained from the different tissues indicated above were fixed in 4% paraformaldehyde for at least 24 h. The tissues were then dehydrated, embedded in paraffin, and cut into 4 μm thick sections using a rotary microtome RM2135 (Leica, Wetzlar, Germany), which were mounted onto gelatincoated glass microscope slides. The sections were stained with hematoxylin and eosin (HE) for histopathological evaluation using standard techniques. Affinity-purified anti-ER and anti-AR primary antibodies were applied at working dilutions of 1:1000 to 1:100 at 4°C for 2 d. Following immunohistochemical staining, the body or tissue sections were counterstained with hematoxylin, dehydrated, and permanently mounted (Permount®, Fisher Scientific, Pittsburgh, PA, U.S.A.) under glass coverslips.
Statistical Analysis
All results are presented as the mean ± SEM (standard error of the mean). The data were analyzed using SPSS for Windows (SPSS Inc., Chicago, IL, USA). A two-way ANOVA was performed using different tissues. When significant differences were obtained between factors, a one-way ANOVA was performed followed by Duncan’s multiple range test. A Student t-test was performed to evaluate the differences between GH/IGF axis genes and MRF genes in each experimental group. Throughout the experiment, probability values lower than 0.05 (P<0.05) and 0.01 (P<0.01) were considered significant.
Results
The expression of ER-β and AR-α1 mRNA in various tissues is presented in Figure 1, showing that both were ubiquitously expressed in the majority of the tissues examined. The expression of ER-β in the liver and gonads of female fish was higher than that of the males in the control group (P<0.05). ER-β expression in the liver of female fish increased significantly following the injection of E2 (P<0.05). Expression of ER-β in the gonad, intestine, and kidney remained unaffected following the injection of sex steroid hormones in both sexes of fish. The hepatic expression of ER-β was higher in the females compared to the males, and an exogenous injection of E2 exhibited an increased expression in the female fish (P<0.05).
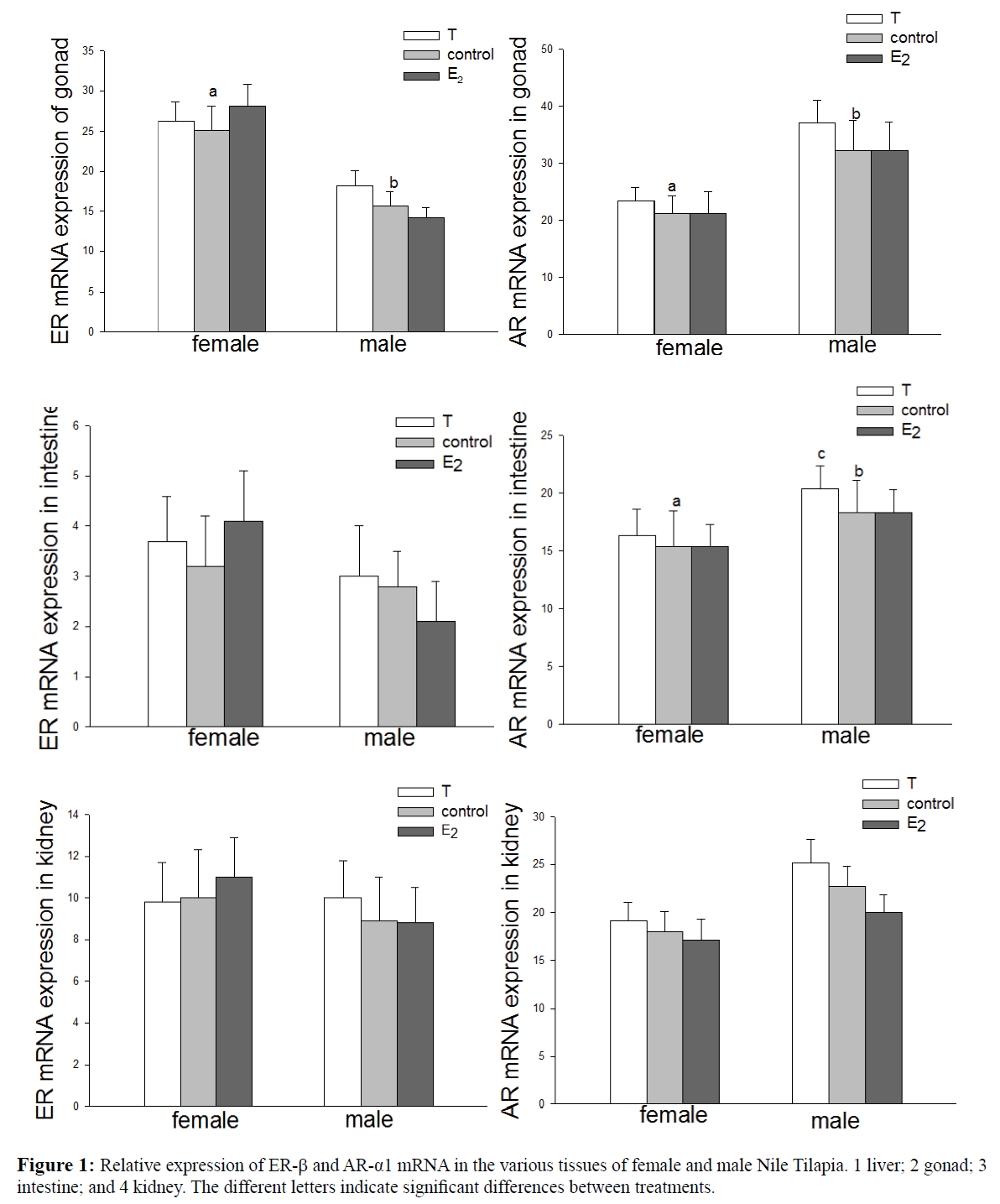
Figure 1: Relative expression of ER-β and AR-α1 mRNA in the various tissues of female and male Nile Tilapia. 1 liver; 2 gonad; 3 intestine; and 4 kidney. The different letters indicate significant differences between treatments.
Expression of ER-β in the gonads was higher in the females than in males (P<0.05). AR-α1 expression was higher in the gonads and intestines of the males compared to the females (P<0.05). An injection of T increased the expression of AR-α1 in the intestines of male fish (P<0.05). Moreover, E2 exhibited no significant influence on the expression of AR-α1 in the liver, gonads, intestines, or kidneys of both male and female fish. A Western blot analysis was conducted and an immunoreactive band is presented in Figure 2. ER was weakly expressed or absent in the liver of the male fish. The ER band in the liver of the female fish was almost 10 times more intensively expressed than in males, and this band was 15% higher in the ovaries than in the testes. In the gonads, the level of AR detected in the males was 25% higher than that in the female fish. In the kidneys, the level of AR detected in the male fish was 50% higher than that in the female fish.
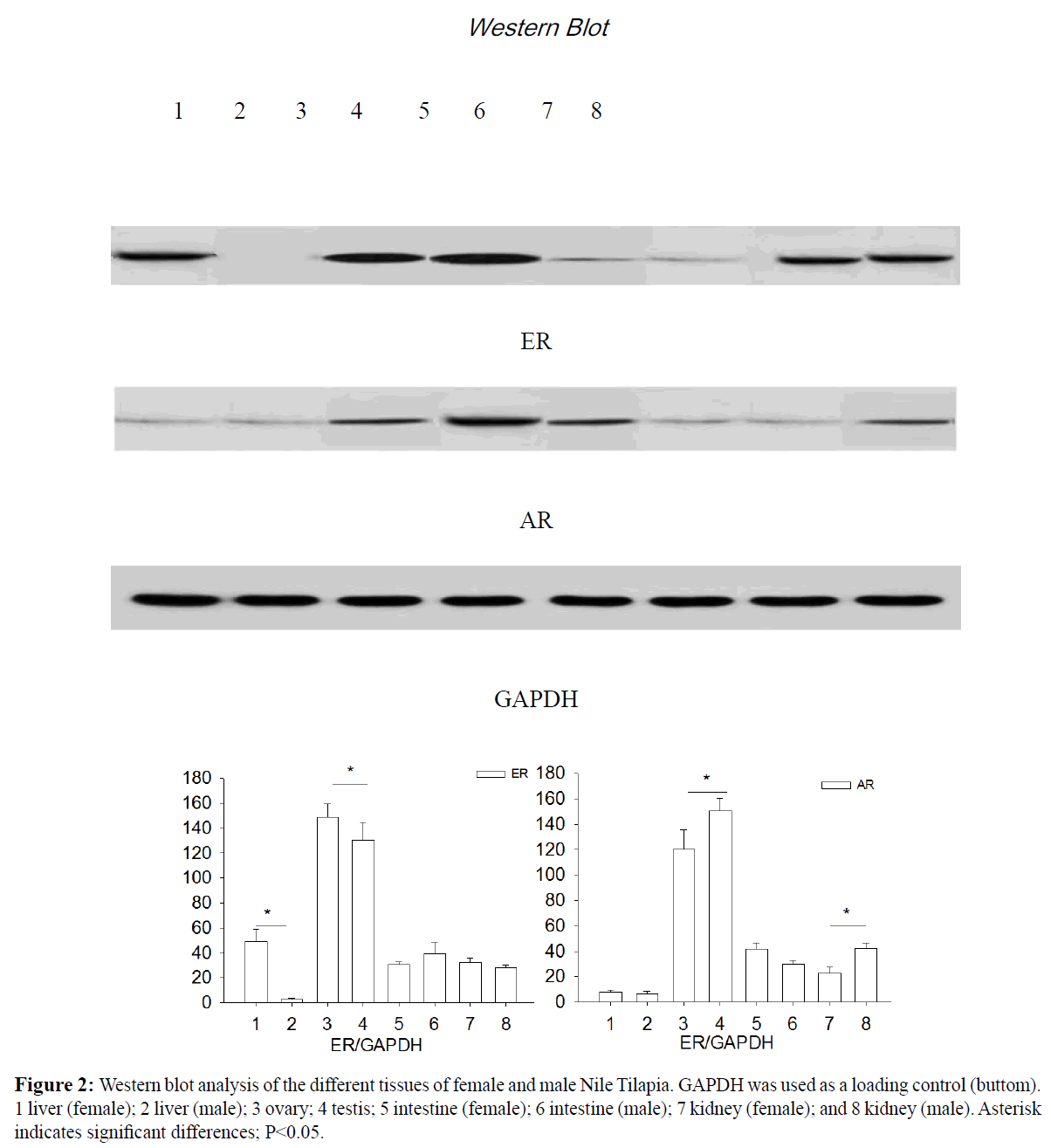
Figure 2: Western blot analysis of the different tissues of female and male Nile Tilapia. GAPDH was used as a loading control (buttom). 1 liver (female); 2 liver (male); 3 ovary; 4 testis; 5 intestine (female); 6 intestine (male); 7 kidney (female); and 8 kidney (male). Asterisk indicates significant differences; P<0.05.
Positive signals of both ER and AR were detected in the gonads, intestines, and kidneys of female and male fish. Weak ER and AR signals were detected in the livers of the female fish (Figure 3E-3P), whereas no signal was detected in the livers of the male fish (Figure 3A-3D). Consistent with the Western blot results, extremely weak and strong ER signals were detected in the testes and ovaries, respectively. In contrast, extremely weak and strong AR signals were detected in the ovaries and testes, respectively (Figure 3F and 3H). At the same time, AR expression was dramatically upregulated in the cells of the intestines of the females compared to the male fish; however, the correlation between sex and ER staining revealed no significant relationship between the two sexes of fish (Figure 3I-3L).
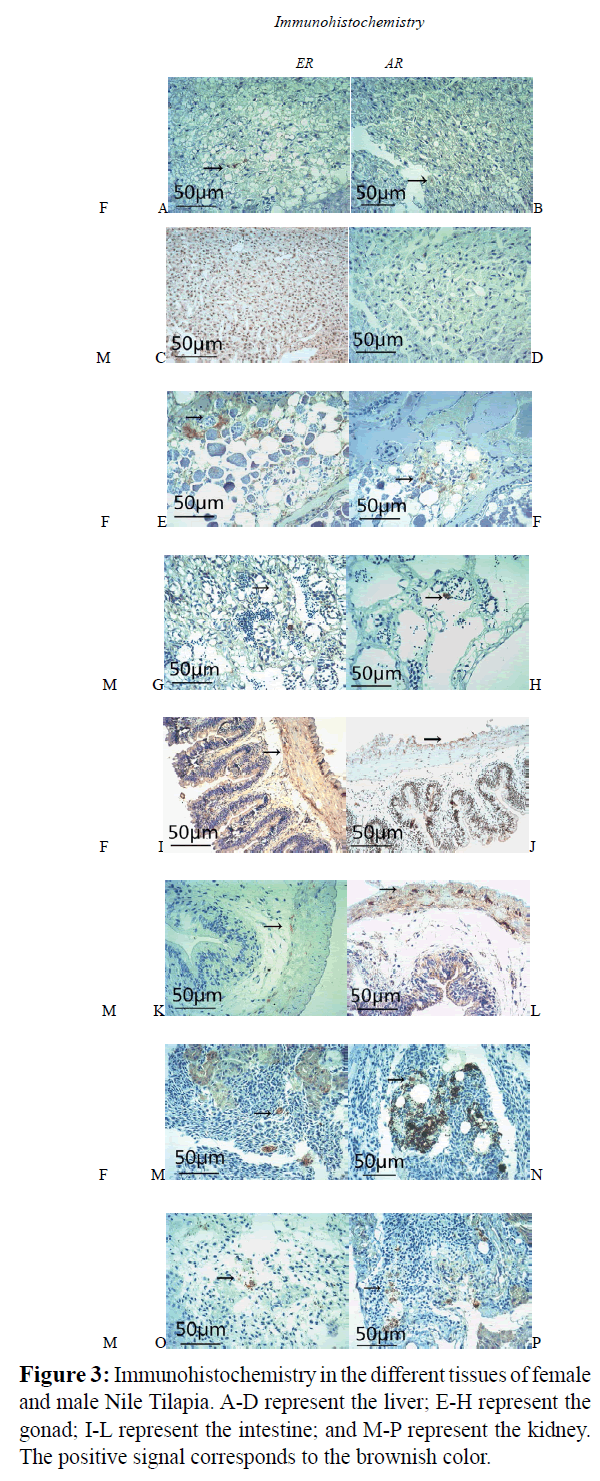
Figure 3: Immunohistochemistry in the different tissues of female and male Nile Tilapia. A-D represent the liver; E-H represent the gonad; I-L represent the intestine; and M-P represent the kidney. The positive signal corresponds to the brownish color.
Discussion
Estrogens are the sex steroid hormones primarily produced by the ovaries through the conversion of cholesterol. Estrogens can also be locally produced in the placenta, adrenal glands, adipose tissue, and brain (Cui, Shen & Li, 2013). Moreover, estrogens are essential for the development and maintenance of the female reproductive, immune, cardiovascular, and peripheral nervous systems (Mauvais-Jarvis, Clegg & Hevener 2013).
Androgens mediate a wide range of male developmental processes, and are particularly important in male sexual differentiation, pubertal maturation, the maintenance of spermatogenesis, and male gonadotropin regulation (Heinlein et al., 2002).
In the present study, the ER was significantly higher in the testes compared to that in the ovaries.Much of the initial research regarding the effects of sex steroids in non-reproductive tissues has focused on the regulation of sex steroids in the liver regarding the growth hormone (GH)/insulin-like growth-factor (IGF) axis, which is central for the systematic regulation of growth in fish (Mccormick, Micheal, Moeckel, Darren & Björnsson 2005; Reinecke M, 2010). It has been shown that E2 increased the growth performance of females in Nile tilapia, while there was no significant effect observed in males. In contrast, T significantly increased the growth performance of males, with no significant effect on females (Yue et al., 2017; Ma, Zhang, Chen & Zhou 2015). In the present study, the level of ER-β mRNA in the liver and gonads of female fish was substantially higher than that of the male fish, which was in accordance with the fact that estradiol regulates the endocrine system of female and male fish. However, the females were found to exhibit more types and a higher expression of ERs (e.g., ER-α, ER-β1, and ER-β2) (Zhang, Huang, Zhu, Liu & Lin 2005). However, according to Zhu, Zhuo, Lu, Huang, Liu & Lin (2008), ER-α1 gradually increased while ER-β1 firstly decreased then positively expressed in testis of male Spinibarbus denticulatus, a local economically important cyprinid fish in China.
Evidence suggests that hepatic vitellogenesis is mediated through both ER- and ER-β (Boyce-Derricott, & Nagler, 2009; Leanos-Castaneda & Van Der Kraak 2007), and the regulation of multiple ER isoforms has been reported in other fish species (Menuet, Le Page, Torres, Kern, Kah & Pakdal 2004; Nelson, Wiehler, Cole & Habibi 2010). In the current study, E2 increased the expression of ER-β mRNA in the liver with no significant effect in the gonads, supporting the role of sex steroids in the regulation of physiological processes directly in the liver during sexual maturation. It should be noted that there was a significant correlation between sex and ER or AR staining in the liver, gonads, and kidneys of tilapia.
In the testes of adult mice, AR was expressed in leydig, sertoli, and most peritubular cells but not in the germ cells, while ER-α was present in leydig and some peritubular cells (Prins, 2002). According to the results of this study, a significant relationship was found between ER and AR expression, as well as sexual growth dimorphism in tilapia. Further research involving a larger series of signaling pathways is recommended, which may be useful for evaluating how sex hormones affect the growth of female and male tilapia.
Acknowledgments
This work was financially supported by SAAS Program for Excellent Research Team (SPERT). Shanghai Municipal Science and Technology Commission (15391912200), Project supported by Shanghai Academy of Agricultural Sciences[2017, A-03], also supported by the China Agriculture Research System (CARS- 49) and the Shanghai Collaborate Innovation Center for Aquatic Animal Genetics and Breeding (ZF1206) to J.L. Zhao.
23063
References
- Phumyu, N., Boonanuntanasarn, S., Jangprai, A. (2012) Pubertal effects of 17α-methyltestosterone on GH-IGF-related genes of the hypothalamic-pituitary-liver-gonadal axis and other biological parameters in male, female and sex-reversed Nile tilapia. General & Comparative Endocrinol 177, 278-292.
- Shupnik, M.A., Rosenzweig, B.A. (1991) Identification of an estrogen-responsive element in the rat LH beta gene. DNA-estrogen receptor interactions and functional analysis. J Biological Chem 266, 17084.
- Heinlein, C.A., Chang, C. (2002) Androgen receptor (AR) coregulators: an overview. Endocrine Reviews 23, 175-200.
- Livak, K.J., Schmittgen, T.D. (2001) Analysis of relative gene expression data using realtime quantitative PCR and the 2−ΔΔCt method. Methods 25, 402-408.
- Kumar, R., Yadav, A., Pakrasi, P.L. (2016) Expression of ER-α and ER-β during peri-implantation period in uterus is essential for implantation and decidualization in golden hamster. Life Sciences 170, 115-122.
- Lee, D.K., Chang, C. (2003) Molecular communication between androgen receptor and general transcription machinery. J Steroid Biochem and Molecular Biol 84, 41-49.
- Smith, L.B., Walker, W.H. (2014) The regulation of spermatogenesis by androgens. Seminars in Cell & Developmental Biology 30, 2-13.
- Prins, G.S. (2002) Localization of androgen and estrogen receptors in adult male mouse reproductive tract. J Andrology, 23, 870-881.
- White, R., Lees, J.A., Needham, M., Ham, J., Parker, M. (1987) Structural organization and expression of the mouse estrogen receptor. Molecular Endocrinol 1, 735.
- Kuiper, G.G., Enmark, E., Pelto-huikko, M., Nilsson, S., Gustafasson, J.A. (1996) Cloning of a Novel Estrogen Receptor Expressed in Rat Prostate and Ovary. Proceedings of the National Academy of Sciences of the United States of America 93, 5925-5930.
- Yue, M.M., Zhao, J.L., Tang S.J. (2017) Effects of Estradiol and Testosterone on the Expression of Growthâ€ÂÂÂÃÂrelated Genes in Female and Male Nile Tilapia, Oreochromis niloticus. J World Aquaculture Society 5.
- Ma, X.L., Zhang, Y., Chen Z.Z., Zhou L.B. (2015) Steroid hormone (E2 and MT) displayed difference in sex for Nile tilapia (Oreochromis niloticus). Oceanologia et Limnologia Sinica 46, 1487-1493.
- Mccormick, S.D., Micheal. F.O., Moeckel. A.M., Darren T.L., Björnsson, B.J. (2005) Endocrine disruption of parr-smolt transformation and seawater tolerance of Atlantic salmon by 4-nonylphenol and 17β-estradiol. General Comparative Endocrinol 142, 280-288.
- Reinecke, M. (2010) Influences of the environment on the endocrine and paracrine fish growth hormone–insulinâ€ÂÂÂÃÂlike growth factorâ€ÂÂÂÃÂI system. J Fish Biology 76, 1233-1254.
- Boyce-Derricott, J., Nagler., J.J. (2009) Regulation of hepatic estrogen receptor isoform mRNA expression in rainbow trout (Oncorhynchus mykiss). General Comparative Endocrinol 161, 73-78.
- Leanos-Castaneda, O., Van Der Kraak. G. (2007) Functional characterization of estrogen receptor subtypes, ERα and ERβ, mediating vitellogenin production in the liver of rainbow trout. Toxicol Applied Pharmacol 224, 116-125.
- Menuet, A., Le Page, Y., Torres, O., Kern, L., Kah, O., et al. (2004) Analysis of the estrogen regulation of the zebrafish estrogen receptor (ER) reveals distinct effects of ERalpha, ERbeta1 and ERbeta2. J Molecular Endocrinol 32, 975-986.
- Nelson, E.R., Wiehler, W.B., Cole, W.C., Habibi, H.R. (2010) Homologous regulation of estrogen receptor subtypes in goldfish (Carassius auratus). Molecular Reproduct Development 74, 1105-1112.
- Cui, J., Shen, Y., Li., R. (2013) Estrogen synthesis and signaling pathways during aging: from periphery to brain. Trends Molecular Medicine 19, 197-209.
- Mauvais-Jarvis, F., Clegg, D.J., Hevener, A.L. (2013) The Role of Estrogens in Control of Energy Balance and Glucose Homeostasis. Endocrine Reviews 34, 309-338.
- Zhang, Y., Huang, W.R., Zhu, P., Liu, X.C., Lin, H.R. (2005) Expression analysis of estrogen related genes in male tilapia. Joint Symposium on the division of Comparative Endocrinology and developmental biology of Chinese Zoological Society. Hangzhou, China 21.
- Zhu, P., Zhang, Y., Zhuo, Q., Lu, D., Huang, J., et al. (2008) Discovery of four estrogen receptors and their expression profiles during testis recrudescence in male Spinibarbus denticulatus. General Comparative Endocrinol 156, 265-276.


