Dalia Ahmed1, Rania Hamed Abdel-Rahman2*, Mohamed Salama3, Laila M El-Zalabany4 and Mona Ahmed El-Harouny4
1Lecturer of Clinical Toxicology and Forensic Medicine, Mansoura University, Mansoura, Egypt
2Assistant Professor of Clinical Toxicology and Forensic Medicine, Mansoura University, Mansoura, Egypt
3Associate professor of Clinical Toxicology and Forensic Medicine, Medical Experimental Research Centre, Mansoura University, Mansoura, Egypt
4Professor of Clinical Toxicology and Forensic Medicine, Mansoura University, Mansoura, Egypt
*Corresponding Author:
Ahmed D
Assistant Professor of Clinical Toxicology and Forensic Medicine
Mansoura University-Faculty of Medicine, Mansoura, Egypt
Tel: +201063831286
E-mail: raniahamed73@gmail.com
Received date: August 03, 2017; Accepted date: August 08, 2017; Published date: August 25, 2017
Citation: Ahmed D, Abdel-Rahman RH, Salama M, El-Zalabany LM, El-Harouny MA. Malathion Neurotoxic Effects on Dopaminergic System in Mice: Role of Inflammation. J Biomedical Sci. 2017, 6(4):30. doi: 10.4172/2254-609X.100074
Copyright: © 2017 Ahmed D, et al. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Keywords
Malathion; Neurotoxicity; Anti-inflammatory drugs
Introduction
Pesticides are utilized for preventing, destroying, repelling or mitigating pests. They are also used in industry, therapeutics and warfare related products [1]. Organophosphate pesticides (OPs) are extensively used worldwide. Their high-intensity occupational application constitutes a global health problem due to the resultant environmental pollution and deleterious chronic health effects especially in developing countries [2,3].
It is worthy-mentioning that prolonged low level OPs exposure is not only limited to occupational settings, but also includes living with those occupationally exposed, living near OPs manufacturing and application sites or ingesting contaminated food [4].
The toxic effects of OPs include different phases i.e. acute toxicity, which involves acute cholinergic crisis due to accumulation of acetyl choline as a result of inhibition of cholinesterase (ChE) enzyme [5]. On the other hand, many reports stated that OPs neurotoxicity (especially in case of chronic exposure) may occur in the absence of ChE inhibition [6-8]. The exact clinical phenotyping of OPs-induced chronic neurotoxicity has been an area of debate with different suggested mechanisms [9,10].
Despite that chronic exposure usually causes symptoms not accompanied by acute cholinergic overstimulation, it is claimed to be associated with neuropsychiatric conditions, such as anxiety and depression. Furthermore, many reports proved positive correlation between different types of OPs and the increased risk of cognitive deficits and neurodegenerative disorders, such as Parkinson’s disease (PD), Alzheimer’s disease, and amyotrophic lateral sclerosis [4,11,12].
Parkinson’s disease is the most common neurodegenerative disorder that is accompanied by motor disability. The hall mark pathological finding in this disease is progressive degeneration of dopaminergic system, mainly the substantia nigra and corpus striatum [13]. The aetiology of sporadic PD (the most common form) is still not fully determined [14]. A current theory assumed that there is a gene-environment interaction, where genetic changes increase the vulnerability of dopaminergic neurons to risk of environmental toxic agents [15].
The mechanism of OPs-induced neurodegeneration is still unclear and seems to be different from AChE inhibition involved in acute toxicity. One suggested mechanism for OPs inducedneurotoxicity is neuroinflammation [9,10]. In the present study, we aimed to verify the degenerative effects of long term exposure to malathion on dopaminergic system in mice and to identify the possible contributing mechanism.
Material and Methods
This study was approved by Medical Research Ethics Committee, Faculty of Medicine, Mansoura University (code no: MD/106). All chemicals were purchased from Sigma -Aldrich™ (Saint Louis, MO, USA) unless otherwise declared.
Animals and Experimental Design
This work was conducted in Medical Experimental Research Centre (MERC), Faculty of Medicine, Mansoura University, Egypt. First, a pilot study was performed to figure out the highest sublethal dose of malathion that produced no toxic clinical cholinergic signs, no morbidities as well as no mortalities. Three doses for malathion were tried (50, 100 and 200 mg/kg given to the mice by gavage).
To conduct the experiment, forty-eight BALB/c adult male mice (Albino inbred strains), and four month-old, 25-30 grams weight, were obtained from MERC. They were housed in clean cages under standard laboratory conditions including suitable temperature (22 ± 2°C), good lighting and aeration. They were fed a standard laboratory diet and tap water.
Mice were randomly divided into four groups (12 mice each) as follows: Control vehicle group (G1) that received distilled water (which is used to dissolve the treatments employed in the current study). Group 2 received malathion in a dose of 200 mg/kg dissolved in distilled water (1/2 ml). Malathion was administered orally by gastric gavage every day for 8 weeks. The other two groups received two different anti-inflammatory drugs (purchased from Amriya Company for pharmaceutical industries) in conjunction with malathion for 8 weeks as follows: Group 3 (G3): "malathion+dexamethasone" M+DX-treated group: mice were given malathion plus subcutaneous dexamethasone injection "3 mg/kg/day" [16] and G4: "malathion+acetyl-salicylic acid" M+ASA-treated group: mice were given malathion plus intraperitoneal injection of ASA "100 mg/kg/day"[17]. Weights of the animals were recorded once a week throughout the experiment.
Assessment of the Neurotoxic Effects of Malathion on Dopaminergic System
Locomotor performance tests
Evaluation of the locomotor activity in mice was done at the end of the 8th week by using ANY-box® (Stoelting Company, USA) which is a multi-configuration behaviour apparatus (Figure 1).

Figure 1: ANY-box ® apparatus (Stoelting Company, USA).
Two different behavioural tests were performed in a room that was completely isolated from external noises:
Parallel rod floor test: Parallel rod floor test apparatus consists of a clear acrylic plastic box 20 cm × 20 cm × 30 cm (width, length, height) and a series of parallel stainless-steel rods placed on stainless steel base plate that acted as a floor for the chamber. The locomotor activity was assessed using two parameters: Foot slips (numbers of errors) which are measured by a touch sensor underneath the parallel rod floor in addition to efficient path (horizontal distance travelled by the mouse in cm).
Open-field test: The apparatus is constructed of a clear acrylic plastic box 40 cm × 40 cm × 35 cm (width, length, height) fits to ANY-box base. Two perpendicular lines were drawn on the floor with a marker and were visible through the clear wall. These lines divided the floor into four equal quadrants: North East (NE), North West (NW), South East (SE) and South West (SW). Each animal was placed individually at the center of the apparatus, allowed to explore it freely and observed for five minutes. Each mouse trial was recorded for latter analysis, using a camera positioned above the apparatus. Locomotor activity was assessed for each mouse using numbers of mid zone cross, immobility, the number of entries into these quadrants and the duration that each animal spent in each quadrant.
Immunohistochemistry
At the end of the second month, the mice were perfused through the aorta with 50 mL of 10 mM phosphate-buffered saline (PBS), followed by 150 mL of a cold fixative consisting of 4% paraformaldehyde, 0.35% glutaraldehyde, and 0.2% picric acid in 100 mM phosphate buffer (PB), under deep anesthesia with pentobarbital (100 mg/kg, i.p.). After perfusion, the brain was quickly removed and post-fixed for 2 days with paraformaldehyde in 100 mM PB and then transferred to 15% sucrose solution in 100 mM PB containing 0.1% sodium azide at 4°C. The brain pieces were processed into paraffin blocks, and then cut by microtome (20 μm). Antigen retrieval was done by emersion of the slides in ethylene diamine tetra-acetic acid (EDTA) solution for 20 minutes in a water bath at 90°C. Slides were then incubated with primary mouse monoclonal antityrosine hydroxylase (TH) antibody (diluted 1:1000) or ionized calcium binding adaptor molecule 1 (anti-iba-1 antibody: diluted 1:2000) over the night at 4°C in a refrigerator. After several washes by PBS, this was followed by biotinylated secondary antibody for 10 min then the avidin-biotin-peroxidase complex (ABC) 10 min at room temperature. All the sections were washed several times with PBS between each incubation, and labeling was then revealed by Diaminobenzidine (DAB) which was used as a chromogen. Slides were counterstained with Meyer’s hematoxylin, dehydrated and cover slipped. Brain sections were examined with standard Olympus® light microscope (model-CX31RTSF).
Image analysis
Analysis was performed by a blinded investigator. The following areas were examined:
Striatal TH-fiber density measurement: Pictures were captured by a digital camera (Olympus® model E-420) and analyzed by using image J software version ij146-jdk6 for windows 7. Mean optical density of TH-positive dopaminergic fibers in the corpus striatum was assessed. To evaluate the entire striatal complex, the images were taken at six rostralcaudal levels corresponding to antero-posterior (AP): +1.60, +1.20, +0.20, -0.30, -0.90 and -1.40 mm from bregma. The striatum in each section was delineated and measured using the image J software. Non-specific background was correlated by subtraction of the non-specific binding as measured from the corpus callosum and for the TH-positive staining completely denervated areas of the striatum. The data represented the average of the six levels [18].
Dopaminergic neurons in Substantia Nigra pars compacta (SNpc): TH-positive cells in the SNpc of both hemispheres were counted, in every fourth section, throughout the entire nucleus. The anatomical levels considered, in the anteroposterior (AP) extension, fell within -5.20 and -5.80 mm, with respect to bregma. Results were expressed as the percentage of cell loss compared to control. Neurodegeneration was ranked from one to four indicating loss of up to 25%, 50%, 75%, 100% in cell count respectively [19].
Microglia in nigrostriatal pathway: Microglial count in SNpc and corpus stiatum was evaluated using anti-Iba1 staining. Cells positive for Iba1 antibodies were counted in the same pattern as TH +ve neurons in both areas.
Statistical Analysis
The statistical analysis of data was performed using the computerized statistical package for the social sciences (SPSS) version 22.0 released 2013 and excel program for figures. Quantitative data was described as mean ± standard error of mean (SEM). For quantitative data; student t-test was used to compare between two groups. One way analysis of variance (ANOVA) test was used to compare more than two groups followed by Tukey’s post hoc test. P is significant if<0.05 at confidence interval 95%.
Results and Discussion
Several researches reported a possible correlation between pesticides exposure and development of neurodegenerative disorders [3,20]. Since OPs are the most widely used pesticides, the previously mentioned studies focused on a link between OPs and PD which is the most common neurodegenerative disease. More interesting, some types of organophosphates have been proved to induce degeneration in the dopaminergic system [10,21].
In the present work, we investigate the neurotoxic effects of chronic malathion administration (an organophosphate compound that is widely used in Egypt) in BALB/c mice behaviourally and pathologically. Open field and parallel rod floor tests were used to assess exploratory behavior, anxiety, locomotor activity and motor incoordination [22].
We found that the change in the weights of the tested animals was insignificant. Some behavioural parameters were significantly affected in the Malathion group in comparison to the control mice. For instance, in parallel rod test, the efficient path was decreased (26.00 ± 2.45) and the number of foot slips was significantly increased in the malathion treated group (3.6 ± 0.24) in comparison with the control group (35.0 ± 1.67 and 0.9 ± 0.28 respectively). With respect to the open field test, the malathion treated mice had more North East entry frequency (2.6 ± 0.40) and they spent less duration in the South East area (27.6 ± 3.72) when compared to the control group (1.3 ± 0.30 and 45.6 ± 6.38 respectively).
It was reported that rats treated with malathion (50 and 100 mg/kg, for 3 days) presented anxiogenic behavior in the openfield test [23]. Additionally, Gould et al. [24] stated that reduced movement around the arena, and reluctance to enter or move from one place to another or spend time in the open central area indicated anxiety-related behaviour in the tested mice as observed in the current study.
It was necessary to verify the pathological impact of malathion on the dopaminergic system of the brains in the studied mice. Tyrosine hydroxylase (TH), an enzyme involved in the synthesis of dopaminergic neurons, was immunehistochemically labelled (TH+) to visualize the dopaminergic neurons and their neuronal processes. Furthermore, ionized calcium binding adaptor molecule 1 (Iba-1), a protein expressed by activated microglia was immunohistochemically labelled to detect microglial cell activation. The observed locomotor deficits were proved to be associated with evident degeneration in the dopaminergic system compared to the control group. Image analysis of corpus striatum showed significant degeneration in G2, treated with Malathion, compared to the control group as illustrated in Figures 2 and 3. The quantification of neurons in substantia nigra revealed parallel loss of dopaminergic neurons. Moreover, quantification of cells positive for Iba1 (microglia) showed accompanying increase in microglial cell count in parallel with microglial activation as evidenced by arborization of cells when examined under high power magnification (4 x) and this also paralleled the resultant neurodegeneration (Table 1 and Figures 2 and 3).
| Groups (n=10 mice each) |
Control group (G1) |
Group 2 |
Group 3 |
Group 4 |
| Parameter |
M (200 mg/kg) |
M (200 mg/kg)
+ Dx (3 mg/kg) |
M (200 mg/kg)
+ ASA (100 mg/kg) |
| (A) Neurobehavioural tests # |
| 1. Parallel rod test |
| Foot slips |
0.9 ± 0.28 |
3.6 ± 0.24 |
1 ± 0.29 |
2.67 ± 0.33 |
| P value |
0.001*a |
0.82a
0.00*b |
0.001*a
0.08b |
| Efficient path |
35.0 ± 1.67 |
26.00 ± 2.45 |
30.00 ± 2.36 |
26.67 ± 1.67 |
| P value |
0.001*a |
0.05*a
0.19b |
0.001*a
0.83b |
| 2. Open field test |
| North East quadrant entry frequency |
1.3 ± 0.30 |
2.6 ± 0.40 |
1.65 ± 0.34 |
2.0 ± 0.17 |
| P value |
|
0.01*a |
0.5a
0.02* b |
0.67a
0.19b |
| South East quadrant stay duration |
45.6 ± 6.38 |
27.6 ± 3.72 |
23.89 ± 7.14 |
22.22 ± 4.8 |
| P value |
|
0.035*a |
0.003*a
0.66b |
0.002*
0.53b |
| Mid zone cross |
1.1 ± 0.23 |
0.80 ± 0.37 |
0.67 ± 0.29 |
1.33 ± 0.29 |
| P value |
|
0.59a |
0.36a
0.81b |
0.62a
0.35b |
| Immobility |
0.3 ± 0.15 |
0.6 ± 0.24 |
0.11 ± 0.11 |
0.00 ± 0.00 |
| P value |
0.06 |
| (B) Immunohistochemical examination of the brains # |
| Fiber density of corpus striatum |
157.92 ± 5.19 |
91.52 ± 1.79 |
127.64 ± 2.58 |
127.12 ± 10.10 |
| P value |
0.001*a |
0.001*a, b |
0.001*a |
| Degree of neuro-degeneration in substantia nigra |
0.00 ± 0.00 |
4.00 ± 0.00 |
1.33 ± 0.14 |
1.58 ± 0.15 |
| P value |
0.001*a |
0.001*a, b |
0.001*a, b |
| Number of activated microglia |
121.33 ± 1.26 |
252.50 ± 19.96 |
187.72 ± 16.79 |
164.91 ± 6.69 |
| P value |
0.001*a |
0.001*a, b |
0.001*a, b |
N.B. M: malathion, ASA: acetyl salicylic acid,
Dx: dexamethasone.
# Data are expressed as mean ± SEM. |
a: means p value compared to the control group whereas
b: means p value compared to M (malathion) group.
*p value is significant if ≤ 0.05.
Table 1: Results of neurobehavioural tests and immunohistochemical examination of the brains in the studied mice groups.

Figure 2: Photomicrographs showing immunohistochemical staining of corpus striatum against anti TH antibody. The photo shows corpus striatum in control group (A), Malathion treated group 200mg/kg (B) and Malathion plus Dexamethasone (3 mg/kg) treated group (C). The photo shows that density of dopaminergic fibers was significantly decreased in Malathion treated group (B) compared to the control (A). However, treatment with anti-inflammatory agent offered neuroprotection to this vulnerablearea (C). (Magnification X4).
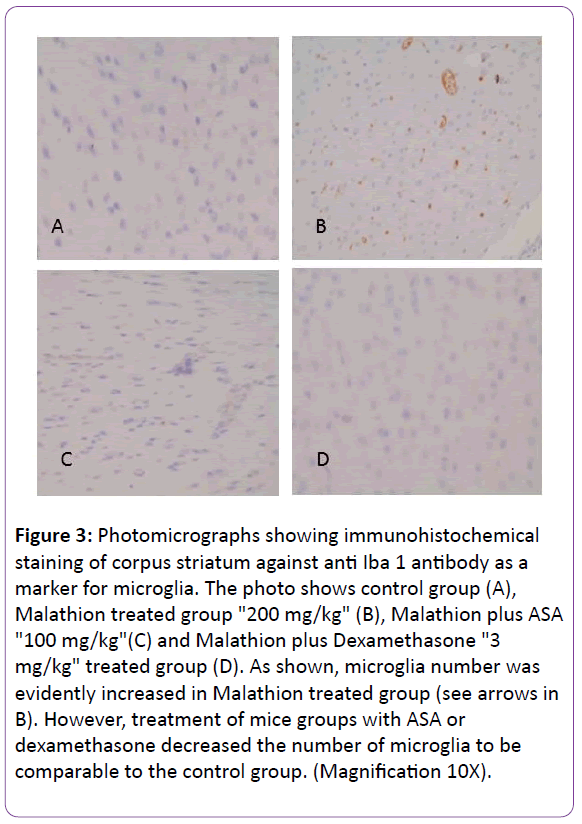
Figure 3: Photomicrographs showing immunohistochemical staining of corpus striatum against anti Iba 1 antibody as a marker for microglia. The photo shows control group (A), Malathion treated group "200 mg/kg" (B), Malathion plus ASA "100 mg/kg"(C) and Malathion plus Dexamethasone "3 mg/kg" treated group (D). As shown, microglia number was evidently increased in Malathion treated group (see arrows in B). However, treatment of mice groups with ASA or dexamethasone decreased the number of microglia to be comparable to the control group. (Magnification 10X).
The results demonstrated in the present research are in accordance with previous findings showing dopaminergic degeneration due to chronic treatment with other types of organophosphates [25-27].
In order to verify the role of OPs induced-neuroinflammation in the current work, we assessed the therapeutic effect of two anti-inflammatory drugs (dexamethasone and ASA) behaviourally and pathologically. Our work revealed better performance in some parameters of the behavioural tests as well as significant improvement of the histopathological findings in G3 & G4 in comparison with the malathion exposed group despite they did not reach the same values of the control mice. Concurrent treatment with dexamethasone or salicylates reduced the number of activated microglia in striata of the treated mice compared to malathion alone treatment. In addition, the brain sections stained against TH showed significant decrease in the number of dopaminergic neurons in the substantia nigra. This cellular loss was parallel with decreased dopaminergic fibres density in corpus striatum. The decrease in microglia, hence neuroinflammation, was also accompanied with significant protection to dopaminergic system as evidenced by less degree of neurodegeneration and cell loss.
Thus, anti-inflammatory drugs administration in conjunction with malathion improved the behavioural dysfunction in malathiontreated mice and was associated with decreased levels of anxiety. Interestingly, anxiety has been reported to be a predominant clinical symptom of neurodegenerative diseases [28]. Therefore, dexamethasone and ASA may have a protective effect in the treatment of neurodegenerative disorderassociated anxiety. Besides, idiopathic PD is characterized by the progressive loss of dopaminergic neurons in the substantia nigra pars compacta leading to dopamine depletion [29] which is also improved after administration of these drugs as demonstrated in the current research.
One important area that correlates OPs and PD is the mechanism of action. Some studies revealed that one of the mechanisms of dopaminergic degeneration is through neuroinflammatory mediators [30,31]. The proinflammatory cytokines and cells play crucial role in the damage of vulnerable dopaminergic cells. On the other hand, several reports showed that OPs induce chronic neurotoxicity through induction of neuroinflammation and could also induce dopaminergic damage through their proinflammatory effects [21,32,33].
In the present work, malathion treated mice showed significant elevation of microglial count and activation in corpus striatum. Worthy mentioning, microglia is the resident immune cells (macrophages) of the brain which can be triggered and activated in response to proinflammatory trigger or neuronal death. Then, several reactive oxygen species and proinflammatory factors (e.g., tumor necrosis factor α, interleukin-1β) are produced and in turn, contributing to neurotoxicity and degeneration [34,35]. In this context, OPs including malathion may initiate an inflammatory response leading to the activation of microglia as noticed in the current work.
These findings denote proinflammatory mechanism of malathion induced neurodegeneration which support previous reports on OPs mechanisms of neurotoxicity and aetiological pathways for dopaminergic system damage [36,37].
One limitation of our study is the lack of assessment of cholinesterase levels. However, it is known that the decreased AChE activity does not correlate with the observed symptoms or reported deficits and manifestations of exposure persist long after ChE levels return to normal [8,38].
In conclusion, the present findings denote the ability of OPs to damage dopaminergic system. Moreover, microglia activation indicates that these effects may be related to malathion induced dopaminergic degeneration and could be induced by neuroinflammation. Targeting inflammation offered neuroprotection in our work, this may advocate the use of antiinflammatory drugs as therapeutic strategy. Further longer studies are warranted to examine the complete reversibility of OPs neurotoxicity in response to different anti-inflammatory agents (the exact doses and duration) and to extrapolate these findings to humans chronically exposed to malathion.
Funding
This research was supported by Medical Experimental Research Centre, Faculty of Medicine, Mansoura University, Egypt.
20190
References
- Mostafalou S, Abdollahi M (2013) Pesticides and human chronic diseases: evidences, mechanisms, and perspectives. Toxicology and Applied Pharmacology 268: 157-177.
- Suratman S, Edwards JW, Babina K (2015) Organophosphate pesticides exposure among farmworkers: pathways and risk of adverse health effects. Rev Environ Health 30: 65-79.
- Sánchez-Santed F, Colomina MT, Hernández EH (2016) Organophosphate pesticide exposure and neurodegeneration. Cortex 74: 417-426.
- Voorhees JR, Rohlman DS, Lein PJ, Pieper AA (2017) Neurotoxicity in Preclinical Models of Occupational Exposure to Organophosphorus Compounds. Frontiers in Neuroscience10: 590.
- Pohanka M (2011) Cholinesterases, a target of pharmacology and toxicology. Biomed. Pap. Med. Fac. Univ. Palacky Olomouc. Czech Repub 155: 219-230.
- Kamel F, Hoppin JA (2004) Association of pesticide exposure with neurologic dysfunction and disease. Environ Health Perspect 112: 950-958.
- Costa LG (2006) Current issues in organophosphate toxicology. Clin Chim Acta 366: 1-13.
- Rohlman DS, Anger WK, Lein PJ (2011) Correlating neurobehavioral performance with biomarkers of organophosphorous pesticide exposure. Neurotoxicology 32:268-276.
- Banks CN, Lein PJ (2012) A Review of Experimental Evidence Linking Neurotoxic Organophosphorus Compounds and Inflammation. Neurotoxicology 33: 575-584.
- Astiz M, Diz-Chaves Y, Garcia-Segura LM (2013) Sub-chronic exposure to the insecticide dimethoate induces a proinflammatory status and enhances the neuroinflammatory response to bacterial lipopolysaccharide in the hippocampus and striatum of male mice. Toxicol. Appl. Pharm 272: 263-271.
- Narayan S, Liew Z, Paul K, Lee PC, Sinsheimer JS, et al. (2013) Household organophosphorus pesticide use and Parkinson’s disease. International Journal of Epidemiology 42: 1476-1485.
- Wang A, Cockburn M, Ly TT (2014) Bronstein, J. and Ritz, B. The Association Between Ambient Exposure to Organophosphates and Parkinson’s Disease Risk. Occup Environ Med 71: 275-281.
- Wirdefeldt K, Adami HO, Cole P (2011) Trichopoulos, D. Mandel, J. Epidemiology and etiology of Parkinson's disease: a review of the evidence. European Journal of Epidemiology 26: 1, S1-S58.
- Cannon JR, Greenamyre JT (2013) Gene-environment interactions in Parkinson’s disease: specific evidence in humans and mammalian models. Neurobiol Dis 57: 38-46.
- Ratner MH, Farb DH, Ozer J, Feldman RG, Durso R (2014) Younger age at onset of sporadic Parkinson's disease among subjects occupationally exposed to metals and pesticides Interdiscip. Toxicol 7: 123-133.
- Drouin-Ouellet J, St-Amour I, Saint-Pierre M, Proulx JL, Kriz J, et al. (2015) Toll-like receptor expression in the blood and brain of patients and a mouse model of Parkinson's disease. Int. J. Neuropsychopharmacol 18: 1-5.
- Thakur P, Nehru B (2013) Anti-inflammatory properties rather than anti-oxidant capability is the major mechanism of neuroprotection by sodium salicylate in a chronic rotenone model of Parkinson’s disease. Neuroscience 231: 420-431.
- Carlsson T, Winkler C, Lundblad M, Cenci MA, Cenci MA, et al. Graft placement and uneven pattern of reinnervation in the striatum is important for development of graft-induced dyskinesia. Neurobiology of Disease 21: 657-668.
- Blandini F, Levandis G, Bazzini E, Nappi G, Armentero MT (2007) Time-course of nigrostriatal damage, basal ganglia metabolic changes and behavioural alterations following intrastriatal injection of 6-hydroxydopamine in the rat: new clues from an old model. Eur. J. Neurosci 25: 397-405.
- Hernández AF, González-Alzaga B, López-Flores I, Lacasaña, M (2016) Systematic reviews on neurodevelopmental and neurodegenerative disorders linked to pesticide exposure: Methodological features and impact on risk assessment. Environ Int 93: 657-679.
- Binukumar BK, Bal A, Gill KD (2011) Chronic dichlorvos exposure: microglial activation, proinflammatory cytokines and damage to nigrostriatal dopaminergic system. NeuroMolecular Medicine 13: 251-265.
- Kamens HM, Crabbe JC (2007) The parallel rod floor test: a measure of ataxia in mice. Nat Protoc 2: 277-281.
- Brocardo PS, Assini F, Franco JL, Pandolfo P, Müller YMR, et al. (2007) Zinc attenuates malathion-induced depressant-like behavior and confers neuroprotection in the rat brain. Toxicol. Sci 97: 140-148.
- Gould TD, Dao DT, Kovacsics CE (2009) In: Mood and Anxiety-related Phenotypes in Mice: Characterization Using Behavioral Tests. ed.Gould T D 1-20.
- Salyha YT (2013) Chlorpyrifos leads to oxidative stress-induced death of hippocampal cells in vitro. Neurophysiol 45: 193-199.
- N’Go PK, Azzaoui F, Ahami AO, Soro PR, Najimi M, et al. (2013) Developmental effects of malathion exposure on locomotor activity and anxiety-like behavior in Wistar rat. Health 5: 603-611.
- Dorri SA, Hosseinzadeh H, Abnous K, Hasani FV, Robati RY, et al. (2015) Involvement of brain?derived neurotrophic factor (BDNF) on malathion induced depressive?like behavior in subacute exposure and protective effects of crocin. Iran J. Basic Med. Sci 18: 958?966.
- Ferretti L, McCurry SM, Logsdon R, McCurry SM, Kukull WA, et al. (2001) Gibbons L and Teri L: Anxiety and Alzheimer's disease. J Geriatr Psychiatry Neurol 14: 52?58.
- Dirnberger G. Jahanshahi M (2013) Executive dysfunction in Parkinson’s disease: A review. Journal of Neuropsychology 7: 193-224.
- Hirsch EC, Vyas S, Hunot S (2012) Neuroinflammation in Parkinson’s disease. Parkinsonism and Related Disorders 18: S210-S212.
- More SV, Kumar H, Kim IS, Song SY, Choi DK (2013) Cellular and molecular mediators of neuroinflammation in the pathogenesis of Parkinson's disease. Mediators Inflamm 13: 12.
- dos Santos AA, Naime AA, de Oliveira J, et al. (2015) Long?term and low?dose malathion exposure causes cognitive impairment in adult mice: evidence of hippocampal mitochondrial dysfunction astrogliosis and apoptotic events. Archives of Toxicology 90: 647–660.
- Callaghan JP, Kelly KA, Locker AR, et al. (2015) Coticosterone primes neuroinflammatory response to DFP in mice: Potential animal model of Gulf War illness. J. Neurochem 133: 708-721.
- McGeer EG, McGeer PL (2010) Neuroinflammation in Alzheimer's disease and mild cognitive impairment: a field in its infancy. J. Alz. Dis 19: 355-361.
- Amor S, Peferoen LA, Vogel DY, Breur M, der Valk PV, et al. (2014) Inflammation in neurodegenerative diseases-an update. Immunology 142: 151-166.
- Torres-Altoro MI, Mathur BN, Drerup JM, Thomas R, Lovinger DM, et al. (2011) Organophosphates dysregulate dopamine signaling glutamatergic neurotransmission and induce neuronal injury markers in striatum. J. Neurochem 119: 303-313.
- Astiz M, Acaz-Fonseca E, Garcia-Segura LM (2014) Sex differences and effects of estrogenic compounds on the expression of inflammatory molecules by astrocytes exposed to the insecticide dimethoate. Neurotox. Res 25: 271-285.
- Rohlman DS, Ismail AA, Rasoul GA, Bonner MR, Hendy O, et al. (2015) A 10-month prospective study of organophosphorus pesticide exposure and neurobehavioral performance among adolescents in Egypt. Elsevier 74: 383-395.

