Key words
Actinomyces hyovaginalis isolate A11-2, Vitamin D3 biotransformation, calcitriol, growth conditions.
Introduction
Vitamin D3 is a fat-soluble prohormone and to exert its function, it must be activated in the liver by 25-hydroxylase to produce 25-hydroxyvitamin D3 (calcidiol). Subsequently, Calcidiol is hydroxylated in the kidney by 1α –hydroxylase to produce the fully active 1α, 25-dihydroxy vitamin D3 (calcitriol). Such hydroxylation steps are mediated by cytochrome P-450 hydroxylase enzymes [1-5]. These biological active forms particularly calcitriol, that exerts the most effect are required for normal mineralization of bone, muscle contraction, nerve conduction, and general cellular function [6-8]. Their deficiency results in impaired bone mineralization, and leads to bone softening diseases such as rickets in children and osteomalacia in adults [9]. However, production of calcitriol is impaired in patients with liver and/or kidney problems. Therefore, synthetic calcitriol has been used clinically in cases of chronic renal failure, hypo-parathyroidism and osteoporosis [10,11]. Unfortunately, the chemical synthesis of calcitriol especially regio- and stereo- selective introduction of a hydroxyl group at C-1 is a very expensive and tedious procedure requiring many reaction steps [12]. The microbial transformation of vitamin D3 has the cost advantages over chemical synthesis however; microorganisms capable of performing this conversion are very limited and belong to the order Actinomycetales particularly the three genera Streptomyces, Amycolataa and Actinomyces [13-15].
In a previous study conducted in our laboratory, a soil isolate, Actinomyces hyovaginalis A11-2 recovered from various soil samples collected from different localities in Egypt was distinguished throughout a screening process by its activity to transform vitamin D3 into calcitriol. This was the first report for vitamin D3 transformation into its biologically active forms by the genus Actinomyces. The present study aimed to study the influence of various culture conditions on vitamin D3 transformation to calcitriol by Actinomyces hyovaginalis isolate A11-2. The studied culture conditions included effect of media composition, different culture media used for preculture, timing of vitamin D3 additions to the main culture, duration of biotransformation process, initial pH, initial quantity of vitamin 3 added, different carbon and nitrogen sources, NaF, dipotassium hydrogen phosphate. Moreover biotransformation of vitamin D3 using cell lysate of the tested isolate was also studied.
Materials and Methods
Microorganism and maintenance condition
Actinomyces hyovaginalis A11-2, an isolate recovered from Egyptian soil. It was maintained onto nutrient agar slants at 4°C. For stock cultures, the isolate cells were concentrated and suspended in slant medium 50:50 and stored at –20oC [16].
Chemicals
Different chemicals used in the present study were of highest quality available and obtained mainly from Sigma-Aldrich (Munich, Germany), El-Nasr chemical Co.(Adwic, Cairo, Egypt) and other local suppliers. While ready made culture media and media ingredients were obtained from Lab M (Topley house, England), Biolife (Milano, Italy), Oxoid (USA), Difco (Detroit, USA) and Life technologies co. (Scotland).
Culture media and biotransformation method.
The basal medium used for vitamin D3 bioconversion consisted of (g/l): defatted soybean, 15; NaCl, 5; CaCO3, 2; K2HPO4, 1; NaF, 0.5 and of pH 7.8. Biotransformation method of vitamin D3 and preparation of the extracts of the biotransformed products were carried out as previously described by Abbas et al., 2011 [15].
Analytical methods
TLC/ HPLC coupled assay. It was used for quantification of the biotransformation product (calcitriol) which was extracted and concentrated as described by Abbas et al., 2011 [15]. The concentrated extract was loaded (40 μl sample) on TLC plastic sheet (Merck, F254 pre-coated) against appropriate amounts of vitamin D3, 1α-hydroxyvitamin D3 and calcitriol standard. Then, the TLC plate was developed ascendingly in a closed glass chamber using chloroform/methanol (10:1 mixture) as a mobile phase. Detection of spots was performed using UV light at 254 nm wavelength. Then, spots obtained on TLC plate, having the same retention factor (Rf) values as calcitriol standard, were scratched from silica gel plates, extracted with methanol of high performance liquid chromatography (HPLC) grade (400 μl) and centrifuged (5000 rpm for 5 minutes). An aliquot (20 μl) of the obtained supernatant was analyzed by HPLC using the following conditions: Zorbax-Agilent C8 column as a stationary phase; methanol/water (90:10) as a mobile phase at flow rate 1 ml/min for 15 min; and UV-VIS detector at wavelength of 254 nm. For exclusion of the interference of calcitriol/vitamin D3 admixture, HPLC analyses were also carried out for the methanolic solutions of scratched TLC spots of calcitriol standard and the mixture of vitamin D3/calcitriol standard. Also, three different concentrations of calcitriol standard were analyzed by HPLC, using the same conditions described of sample analysis. The average of the peak areas equivalent to the concentrations applied was calculated and used to calculate the amount of the produced calcitriol in samples [15]. At least 3 HPLC runs were made per parameter test under the same conditions.
Optimization of vitamin D3 biotransformation
Effect of different culture media used for preculture. Nutrient broth, originally used as the preculture medium, was replaced with different media which included sabauroud dextrose, starch nitrate and YMG (consisted of (g/l): yeast extract, 4 g; malt extract, 10 g; glucose, 4 g; pH 7.2) broths. Other growth conditions for both the preculture and the main culture were kept constant and carried out as follows. A single colony of the bacterial isolate was transferred to 10 ml nutrient broth (preculture) contained in 100-ml Erlenmeyer flask which was incubated in shaking incubator at 28°C and 200 rpm for 2 days. An aliqout (1 ml) of the preculture obtained was used to inoculate 50 ml basal medium used for vitamin D3 bioconversion contained in 250-ml Erlenmeyer flask (main culture), incubated at 28°C and 200 rpm for 2 days. Thereafter, 10 mg of vitamin D3 (dissolved in 250 μl 96 % ethanol) was added and the culture was continued, under the same conditions, for another 7 days [15].
Effect of timing of vitamin D3 addition. Basically, Vitamin D3 substrate was added 2 days after the beginning of the main culture, other addition times were tried: 0 day (simultaneous with the inoculum), 1 day and 3 days after inoculation and incubation. Preculture (using nutrient broth), main culture and culture conditions were as previously mentioned.
Effect of duration of the bioconversion process. Basically, the biotransformation reaction was continued for 7 days. In this protocol, different periods of duration were tried: 48, 60, 72, 84, 96, 120 and 144 hrs. The culture conditions and the process were completed as previously described.
Effect of initial pH of basal medium. This was carried out by conducting the biotransformation reaction in the basal medium at different initial pH values (6, 7, 7.5 and 8.4) around the pH 7.8 that was originally applied. The culture conditions and the process were completed as previously described except that the main culture was incubated for 4 days only after addition of vitamin D3.
Effect of the initial quantity of added vitamin D3. Different initial amounts (2; 10; 40 or 100 mg% w/v) around the originally tested amount (20 mg% w/v) were used and their effect on the biotransformation efficiency was determined. The culture conditions and the process were completed as previously described except that the main culture was incubated for 4 days only after addition of vitamin D3.
Effect of different basal medium ingredients
Effect of separate removal of some basal medium ingredients. The basal medium, that was applied previously for vitamin D3 biotransformation, was deprived of one of its ingredients in a time except that both sodium chloride and calcium carbonate were always maintained at their original concentrations. This was done to determine which component(s) is/are crucial for the biotransformation process. Media deprived each of glucose, defatted soyabean, dipotassium hydrogen phosphate or sodium fluoride were formulated and designated M1, M2, M3 and M4, respectively. The culture conditions and the process were completed as previously described except that the main culture was incubated for 4 days only after addition of vitamin D3 .
Effect of carbon source and its concentration. Glucose of basal medium was replaced with different carbon sources which included fructose, galactose, lactose, sucrose, malt extract, soluble starch and sodium citrate. These carbon sources were tested separately at the same applied concentration of glucose (1.5% w/v). The carbon source(s) proved to be optimum for calcitriol production was/were further tested at different concentrations. The process were completed as previously described except that the main culture was incubated for 4 days only after addition of vitamin D3.
Effect of nitrogen source and its concentration. Defatted soyabean of basal medium was replaced with different nitrogen sources which included corn oil, corn steep liquor, yeast extract, cooked meat, skim milk and diammonium hydrogen phosphate. These nitrogen sources were tested separately at the same applied concentration of defatted soyabean (1.5% w/v). The nitrogen source(s) proved to be optimum for calcitriol production was/were further tested at different concentrations. The culture conditions and the process were completed as previously described except that the main culture was incubated for 4 days only after addition of vitamin D3.
Effect of different concentrations of sodium fluoride and dipotassium hydrogen phosphate. Different concentrations of sodium fluoride (20, 40, 50 (basal used concentration), 100 and 200 mg% w/v) were investigated. For dipotassium hydrogen phosphate, the investigated concentrations included 20, 40, 100 (basal used concentration), 200 and 2000 mg% w/v. For all concentrations tested, initial pH was adjusted to that of the basal medium (pH 7.8). The culture conditions and the process were completed as previously described except that the main culture was incubated for 4 days only after addition of vitamin D3.
Effect of incorporation of some dispersing agents and surfactants. The dispersing agents tested included polyethylene glycol 400 (PEG-400) and propylene glycol. Tween 80 was a representative of surfactants. Each of the tested agents was added to the main culture, at 0.5% v/v, simultaneously with vitamin D3 addition. The culture conditions and the process were completed as previously described except that the main culture was incubated for 4 days only after addition of vitamin D3.
Testing vitamin D3 biotransformation in two modified media. From the different experiments previously conducted, the medium ingredients proved to be optimum for vitamin D3 biotransformation by the test isolate were collected together in two different formulated media, SkM and SbM (Table 1) and the results were compared to that of basal medium. The culture conditions and the process were completed as previously described except that the main culture was incubated for 4 days only after addition of vitamin D3.
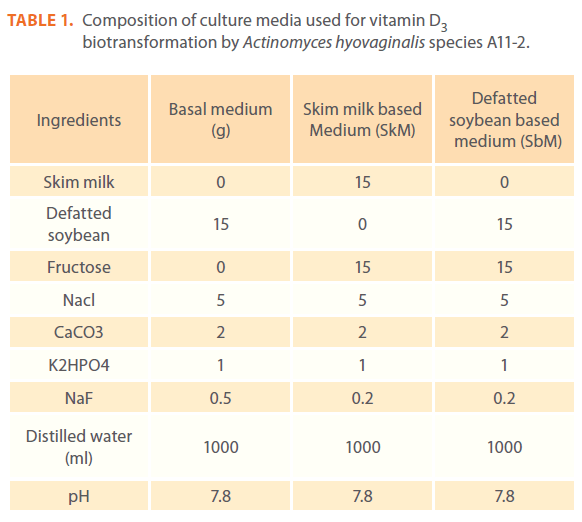
Table 1. Composition of culture media used for vitamin D3 biotransformation by Actinomyces hyovaginalis species A11-2.
Biotransformation of vitamin D3 using cell lysate of the test isolate
Preparation of cell lysate of the test isolate. Preculture, main culture and culture conditions were as previously mentioned except that after 2 days incubation of inoculated main culture, the cells were harvested instead of adding vitamin D3 at that time and continuing incubation. The harvested cells were washed three times with and re-suspended in 10 ml of medium consisting of: NH4)2HPO4 (0.2 M), 45 ml; NH4H2PO4 (0.2 M), 40 ml; NaCl, 1 g; NaF, 0.1 g; K2HPO4, 0.2 g; MgSO4.7H2O, 0.1 g; distilled water to 115 ml; pH 7.8. The suspended cells were then sonicated in 50-ml beaker, kept in ice, at 70% power for 3 successive times; each for 1 min with 30 seconds intervals.
Vitamin D3 biotransformation procedure using the prepared cell lysate. An amount (10 mg) of vitamin D3 dissolved in 250 μl 96 % ethanol, was added to the flask containing the sonicated biomass prepared above. The flask was then incubated in shaking incubator at 28°C and 200 rpm for 6 hrs. After that, the flask contents were extracted and analyzed as previously described. The effect of using different cell counts for the preparation of cell lysate of the test isolate on the biotransformation of vitamin D3 was also investigated.
Results
HPLC analyses of TLC spot of vitamin D3 biotransformation product.
HPLC analyses of the methanolic extracts of the produced TLC spots of isolate A11-2 (the spot with Rf comparable to that of calcitriol), calcitriol standard and mixture of calcitriol standard/ vitamin D3 were carried out and the results are shown in Figure 1.

Figure 1: HPLC analyses of the methanolic extracts of TLC spots of isolate A11-2, the spot with Rf comparable to that of calcitriol, (a); calcitriol standard, (b) and mixture of calcitriol standard/vitamin D3, (c) Retention times are shown between brackets.
Optimization of vitamin D3 biotransformation
It was found that the treatment of growth supernatant of the test isolate with ammonium sulphate before the extraction process had a negative effect on calcitriol recovery where lower calcitriol amount (about 45% reduction compared to that obtained without treatment by ammonium sulphate) was obtained.
Effect of different culture media used for preculture. As shown in Figure 2, nutrient broth gave the highest amount of calcitriol followed by starch nitrate broth then YMG broth. Regarding growth, both nutrient broth and YMG broth produced comparable viable cell counts which were higher than that obtained with starch nitrate broth. On the other hand, sabouraud dextrose broth did not support the growth of the test isolate and consequently no calcitriol production occurred.

Figure 2: Effect of different culture media used for preculture on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2.
Effect of timing of vitamin D3 addition. As shown in Figure 3, addition of vitamin D3 two days after beginning of the main culture gave the highest amount of calcitriol. Whereas, low amounts of calcitriol were obtained with other addition times. Regarding growth, the highest viable cells count was obtained when vitamin D3 was added three days after beginning of the main culture.

Figure 3: Effect of timing of vitamin D3 addition to the main culture on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2.
Effect of initial quantity added of vitamin D3. As shown in Figure 4, maximum calcitriol production was achieved upon addition of vitamin D3 at 20 mg% w/v. Regarding growth, nearly no pronounced change in viable cells count was obtained at the different quantities of added vitamin D3 (2; 10; 20; 40 and 100 mg% w/v).

Figure 4: Effect of initial quantity of vitamin D3 added to the main culture on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2.
Effect of duration of the bioconversion process. As shown in Figure 5, continuing the main culture for 96 hrs after addition of vitamin D3 gave the highest amount of calcitriol. Whereas, other tested duration times produced low amounts of calcitriol with different degrees. Regarding growth, duration times of 72, 84 and 96 hours produced comparable viable cells counts higher than those produced by other tested duration times. According to the obtained results, the duration times of 96 hrs was applied in subsequent experiments.

Figure 5: Effect of duration time of vitamin D3 bioconversion reaction on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2.
Effect of Initial pH of basal medium. Testing the bioconversion of vitamin D3 and calcitriol production by the test isolate at different initial pH values (6, 7, 7.1, 7.5, 7.8 and 8.4) showed maximum calcitriol production at initial pH 7.8 (Figure 6). Regarding growth, comparable viable cells counts were obtained over the pH range of 7-7.8 while the initial pH values of 6 and 8.4 caused pronounced reduction of growth.

Figure 6: Effect of initial pH value of main culture medium on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2.
Effect of different basal medium ingredients
Effect of separate removal of some basal medium ingredients. As shown in Figure 7, using the complete basal medium for main culture gave the highest amount of calcitriol. Whereas, deprivation of the basal medium each of glucose, defatted soyabean, sodium fluoride or dipotassium hydrogen phosphate (formulated media M1, M2, M3 and M4, respectively) produced low amounts of calcitriol. The growth patterns results revealed maximum growth with M3 medium.

Figure 7: Effect of separate removal of some basal medium ingredients on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2. M1, M2, M3 and M4 are basal medium deprived of glucose, defatted soyabean, dipotassium hydrogen phosphate and sodium fluoride, respectively.
Effect of replacement of basal medium glucose with other carbon sources. Replacing basal medium glucose with other carbon sources (including fructose, galactose, lactose, sucrose, malt extract, soluble starch and sodium citrate), at the same glucose concentration applied (1.5% w/v) showed different levels of calcitriol production and also different viable cells counts (Figure 8). The highest calcitriol production was obtained with fructose while the highest viable cells count was obtained with sucrose.

Figure 8: Effect of replacement of basal medium glucose with other carbon sources on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2. The tested carbon source was applied at the same concentration of glucose originally present in basal medium (1.5% w/v).
Effect of different concentrations of glucose and fructose. From the results obtained before, it was found that both glucose and fructose gave higher calcitriol production by the test isolate as compared to other tested carbon sources. Accordingly, these two carbon sources were further examined at different concentrations (0.5, 1, 1.5, 2 and 2.5% w/v). The results, shown in Figure 9 a and b, revealed that the maximum calcitriol production was achieved at 1.5% w/v concentration for glucose or fructose. For both tested sugars, high growth was attained at high sugar concentrations (1.5, 2 and 2.5% w/v).

Figure 9: Effect of different concentrations of glucose (a) and fructose (b) in main culture medium on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2.
Effect of replacement of basal medium defatted soyabean with other nitrogen sources. Replacing basal medium defatted soyabean with other nitrogen sources (including corn oil, corn steep liquor, yeast extract, cooked meat, skim milk and diammonium hydrogen phosphate), at the same defatted soyabean concentration applied (1.5% w/v) showed different levels of calcitriol production and also different viable cells counts (Figure 10). The highest calcitriol production and the highest viable cells count were both obtained with skim milk.

Figure 10: Effect of replacement of basal medium defatted soyabean with other nitrogen sources on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2. The tested nitrogen source was applied at the same concentration of defatted soyabean originally present in basal medium (1.5% w/v).
Effect of different concentrations of defatted soyabean and skim milk. From the results obtained before, it was found that both defatted soyabean and skim milk gave higher calcitriol production by the test isolate as compared to other tested nitrogen sources. Accordingly, these two nitrogen sources were further examined at different concentrations (0.5, 1, 1.5, 2 and 2.5% w/v). The results, shown in Figure 11 a and b, revealed that the maximum calcitriol production was achieved at 1.5% w/v concentration for defatted soyabean or skim milk. For both tested nitrogen sources, high growth was attained at high concentrations (1.5, 2 and 2.5% w/v).
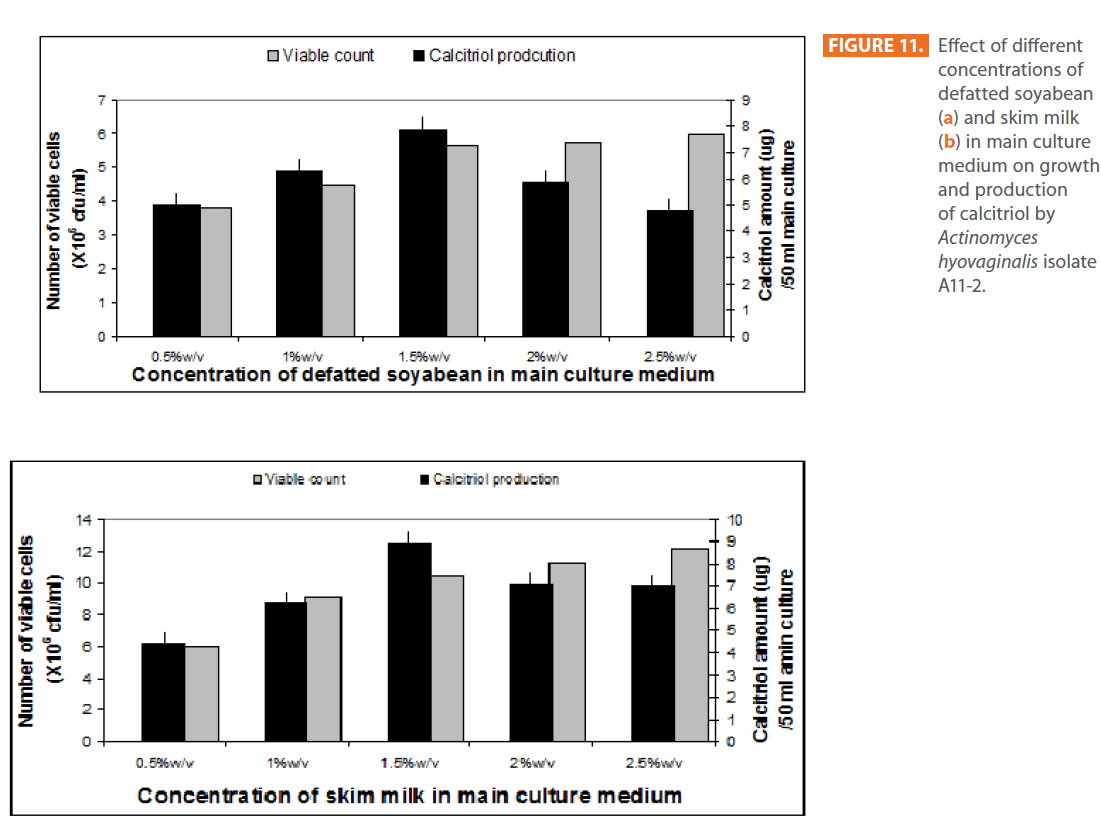
Figure 11: Effect of different concentrations of defatted soyabean (a) and skim milk (b) in main culture medium on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2.
Effect of different concentrations of sodium fluoride. Testing the effect of different concentrations of sodium fluoride, below and above the concentration originally present in basal medium (50 mg% w/v), showed an increase in both calcitriol production and growth at the concentration of 20 mg% w/v as compared to other tested concentrations (Figure 12).

Figure 12: Effect of different concentrations of sodium fluoride in main culture medium on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2.
Effect of different concentrations of dipotassium hydrogen phosphate. Testing the effect of different concentrations of dipotassium hydrogen phosphate, below and above the concentration originally present in basal medium (100 mg% w/v), showed the maximum calcitriol production at the concentration of 100 mg% w/v. whereas, the highest growth was attained at the concentration of 20 mg% w/v (Figure 13).

Figure 13: Effect of different concentrations of dipotassium hydrogen phosphate in main culture medium on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2. For all concentrations tested, initial pH was adjusted to that of the basal medium (pH 7.8).
Effect of incorporation of propylene glycol, polyethylene glycol 400 and tween 80. As shown in Figure 14, incorporating each of propylene glycol, polyethylene glycol 400 or tween 80 to the main culture medium (simultaneously with vitamin D3 addition and at concentration of 0.5% v/v) affected both calcitriol production and growth negatively.

Figure 14: Effect of incorporation of propylene glycol, polyethylene glycol 400 and tween 80 to the main culture medium on growth and production of calcitriol by Actinomyces hyovaginalis isolate A11-2.
Testing vitamin D3 biotransformation in two modified media. As shown in Figure 15, the wild type Actinomyces hyovaginalis isolate A11-2 showed the highest growth in SbM medium. However, it showed maximum calcitriol production in SkM.

Figure 15: Growth and vitamin D3 biotransformation by Actinomyces hyovaginalis isolate A11-2 in two modified main culture media (SkM and SbM)
Biotransformation of vitamin D3 using cell lysate of the test isolate. The results revealed that a cell lysate (prepared from 1.5X107 cfu) produced 5.9 μg calcitriol (in 6 hrs) while the intact cells (of 2.6X108 cfu count) of the test isolate produced calcitriol amount of 8 μg in 96 hrs (Table 2). The same experiment was repeated several times using different cell counts for the preparation of cell lysate of the test isolate to determine the effect of the biomass on the biotransformation of vitamin D3. Results revealed that as the initial cell count used to prepare the cell lysate increased, the amount of calcitriol produced increased (data not shown).

Table 2. Biotransformation of vitamin D3 into calcitriol using intact cells and cell lysate of Actinomyces hyovaginalis isolate A11-2.
Discussion
TLC/HPLC coupled assay was used to quantitate the production of calcitriol by Actinomyces hyovaginalis isolate A11-2 during the physiological optimization. Under the applied HPLC conditions and for calcitriol standard/vitamin D3 mixture, the retention time of both calcitriol standard and vitamin D3 were 1.7 and 11 min, respectively. The 1.7 min retention time was also obtained when each of calcitriol standard and the sample (methanolic solution of scratched spot with Rf value of 0.25 on TLC plate) were analyzed by HPLC (Figure 1). This gives an additional evidence for the identity of calcitriol as a biotransformation product by Actinomyces hyovaginalis isolate A11-2.
Optimization of vitamin D3 biotransformation
Treatment of growth supernatant before product extraction of the test isolate with ammonium sulphate had a negative effect on calcitriol recovery where lower amount of calcitriol (about 45% reduction compared to that obtained without treatment by ammonium sulphate) was obtained. The possible reason for that was either the interference of ammonium sulphate with the subsequent extraction process of the formed calcitriol and/ or the adsorption of the formed calcitriol on the precipitated proteins. Accordingly, no treatment with ammonium sulphate was carried out for calcitriol recovery from the growth supernatant of the test isolate in subsequent experiments.
Effect of preculture medium, timing and quantity of added vitamin D3, duration of the bioconversion process and Initial pH of basal medium. Different preculture media used for inoculum preparation of the test isolate were tested for their effect on vitamin D3 biotransformation. These media included nutrient, starch nitrate, YMG and Sabauroud’s dextrose broths. The results in (Figure 2) revealed the absence of coherency between the effect of preculture on growth and its effect on calcitriol production. This was shown by that while both nutrient and YMG broths gave the same amount of growth, they affected vitamin D3biotransformation differently. The test isolate showed higher calcitriol production with nutrient broth. In the same way, although both starch nitrate and YMG broths gave different amounts of growth, yet they exhibited similar amounts of calcitriol. Sabauroud’s dextrose broth supported neither growth nor calcitriol production. Accordingly, the preculture type for inoculum preparation affects the biotranformation capability of the Actinomyces hyovaginalis isolate A11-2 for vitamin D3 and this should be considered for industrial application. Similarly, such effect was found in other biotransformation processes [17].
Addition of vitamin D3 two days after beginning of main culture was found to be the optimal timing for the highest production of calcitriol (Figure 3). These findings differ from those seen in Kang’s work [18] for optimization of vitamin D3biotransformation into calcitriol where his group found that simultaneous addition of vitamin D3, with the inoculation of the main culture, caused the highest production of calcitriol. Increasing the quantity of vitamin D3, added to the main culture, from 1 mg to 10 mg resulted in accompanied rise in the production of calcitriol though the number of viable bacterial cells remained nearly constant (Figure 4). However, using the substrate at initial quantities higher than 10 mg (20 mg and 50 mg) led to slight decline in the number of viable bacterial cells and sharp decrease in the quantity of the product which might be attributed to the inhibitory effect of vitamin D3 excess on the bioconversion process.
Increasing the duration of the bioconversion process resulted in a rise in viable count of Actinomyces hyovaginalis isolate A11-2 with concomitant increase in the production of calcitriol reaching its maximum value when the reaction was continued up to 96 hrs followed by a substantial decrease thereafter (Figure 5). Accordingly, the duration time of 96 hrs was applied in subsequent experiments. Viable biomass showed the highest values over the duration times 72-120 hrs. This finding verified that the duration of contact of the bacterial cells with the substrate is an important factor affecting the bionconversion process. However, the decline in the production of calcitriol after contact time of 96 hrs could be attributed to the degradation of the formed calcitriol. These findings differ from those seen in Kang’s work for optimization of vitamin D3biotransformation into calcitriol where his group found that duration of 168 hrs caused the highest production of calcitriol [18].
Increasing the initial pH from 6 to pH 7.8 caused a gradual increase in the production of calcitriol with a marked increase at pH 7.8. Further increase of pH to 8.4 caused a sharp decrease in calcitriol production (Figure 6). This finding is in accordance with that obtained by Kang et al., 2006 [18]. They found that the bioconversion process is affected by pH, recording highest value in slightly alkaline pH range. Regarding growth, comparable viable cells counts were obtained over the pH range of 7-7.8 while the initial pH values of 6 and 8.4 caused pronounced reduction of growth.
Effect of different basal medium ingredients
Effect of separate removal of some basal medium ingredients. Glucose, defatted soyabean, sodium fluoride and dipotassium hydrogen phosphate were found to be crucial for the bioconversion process of vitamin D3 into calcitriol by Actinomyces hyovaginalis isolate A11-2 where their separate removal caused marked reduction in calcitriol production (Figure 7). It was found that calcitriol production was declined to nearly half its value by removing each of glucose, defatted soyabean or sodium fluoride. Whereas, removal of dipotassium hydrogen phosphate decreased the production of calcitriol to quarter its value. Surprisingly, the lowest calcitriol production achieved with dipotassium hydrogen phosphate removal was accompanied with the highest biomass production. Removal of other tested ingredients caused decrease in growth with different degrees.
Effect of carbon source and its concentration. In order to determine the effect of the carbon source type on calcitriol production, glucose of the basal medium was replaced with various carbon sources. Using fructose as a substitute for glucose in the main culture medium caused the highest production of calcitriol (1.6 fold increase in production as compared to glucose) (Figure 8). This finding differs from that seen in Kang’s work for optimization of vitamin D3 biotransformation into calcitriol where his group found that using glucose as the carbon source caused the highest calcitriol production while using fructose as a substitute for glucose decreased calcitriol production sharply [18]. Other carbon source substitutes (galactose, lactose, sucrose, malt extract, soluble starch and sodium citrate) did not increase calcitriol production compared to glucose. Although sucrose caused the maximum viable cells count among all tested carbon sources. Yet, its use decreased the production of calcitriol to 75% as compared to that produced with glucose. In addition, other carbon sources, except fructose, caused decrease of the growth as compared to glucose.
To study the effect of carbon source concentration on the bioconversion process, two carbon sources were selected which included glusoce (basal medium carbon source) and fructose (which caused the highest calcitriol production). Incorporation of either glucose or fructose in the main culture medium at concentration of 1.5%w/v led to the highest production of calcitriol by the test isolate. Whereas, concentrations below or above this value of either sugar decreased calcitriol production (Figure 9 a & b). This reduction indicates that sugar concentrations higher than 1.5% w/v have inhibitory effect on calcitriol production. For both sugars, number of viable cells increased linearly with the increase of the applied concentration up to 1.5% w/v followed by no appreciable increase in viable cell count.
Effect of nitrogen source and its concentration. In order to determine the effects of the nitrogen source on calcitriol production, defatted soyabean of the basal medium was replaced with various nitrogen sources. Out of the nitrogen sources investigated, skim milk was found to be the most effective for calcitriol production (1.3 fold increase in production as compared to defatted soyabean) (Figure 10). This finding is similar to that seen in Kang’s work for optimization of vitamin D3 biotransformation into calcitriol. In all cases, calcitriol production was found to be related to the number of viable cells for the tested nitrogen sources [18].
To study the effect of nitrogen source concentration on the bioconversion process, two nitrogen sources were selected which included defatted soyabean (basal medium nitrogen source) and skim milk (which caused highest calcitriol production). Incorporation of either defatted soyabean or skim milk in the main culture medium at different concentrations (0.5, 1, 1.5, 2 and 2.5% w/v) caused a gradual increase of both growth and calcitriol production up to 1.5%w/v followed by a plateau in case of growth and a little decrease in calcitriol production (Figure 11 a & b).
By comparing the results in Figures 10 and 12, it is shown that the profiles of growth and calcitriol production in relation to concentrations applied of carbon sources (glucose and fructose) and nitrogen sources (defatted soyabean and skim milk) have high similarity. Both carbon and nitrogen sources showed an increase in growth and calcitriol productin by increasing the concentration applied up to 1.5% w/v in all cases. Further increase in concentrations had nearly no effect on growth while calcitriol production showed a decrease which was more apparent in case of carbon sources especially fructose.
Effect of different concentrations of sodium fluoride and dipotassium hydrogen phosphate. Testing the effect of different concentrations of sodium fluoride, below and above the concentration originally present in basal medium (50 mg% w/v), showed an increase in both calcitriol production and growth at the concentration of 20 mg% w/v as compared to other tested concentrations (Figure 12). Calcitriol production profile showed a gradual decrease by increasing sodium fluoride concentration up to 100 mg% and no further decrease at 200 mg%. Whereas, growth exhibited the same gradual decrease by increasing sodium fluoride concentration but along all the concentrations applied.
Testing the effect of different concentrations of dipotassium hydrogen phosphate, below and above the concentration originally present in basal medium (100 mg% w/v), showed the maximum calcitriol production at the concentration of 100 mg% w/v. whereas, the highest growth was attained at the concentration of 20 mg% w/v (Figure 13). In contrast to carbon sources (glucose and fructose), nitrogen sources (defatted soyabean and skim milk) and sodium fluoride, growth and calcitriol production in relation to concentration in case of dipotassium hydrogen phosphate showed different profiles. Calcitriol production profile showed an increase by increasing dipotassium hydrogen phosphate concentration up to 100 mg% followed by gradual decrease at higher applied concentrations. On the other hand, maximum growth was obtained at the lowest dipotassium hydrogen phosphate concentration applied (20 mg%) and a gradual pronounced decrease was obtained at higher concentrations. The finding obtained in the present study indicated that dipotassium hydrogen phosphate is important for the bioconversion process but this effect is concentration dependent [18].
Effect of incorporation of propylene glycol, polyethylene glycol 400 and tween 80. As shown in Figure 14, incorporation each of propylene glycol, polyethylene glycol 400 and tween 80 to the main culture medium (simultaneously with vitamin D3 addition and at concentration of 0.5% v/v) affected both growth and calcitriol production negatively. Addition of propylene glycol led to slight decline in the viable count but significant decrease in calcitriol production. This finding is similar to that seen in Kang’s study for optimization of vitamin D3 biotransformation into calcitriol [18]. Although the incorporation of PEG 400 decreased the viable cells count to a lower degree than that caused by propylene glycol, it caused a less reduction in calcitriol production as compared to propylene glycol. Addition of tween 80 sharply decreased both growth and calcitriol production.
Testing vitamin D3 biotransformation in two modified media. The two culture media (SkM and SbM) were similarly formulated from the ingredients and at the concentrations that showed maximum calcitriol production except that SkM contained skim milk as nitrogen source while SbM contained defatted soyabean as nitrogen source. As shown in Figure 15, the Actinomyces hyovaginalis isolate A11-2 showed the highest growth in SbM medium. However, it showed maximum calcitriol production in SkM. The coherency between growth and calcitriol production could not be established.
Biotransformation of vitamin D3 using cell lysate of the test isolate. This experiment was conducted to test vitamin D3 biotransformation using cell lysate of the test isolate. The result was compared to that obtained using intact cells remained under the growth conditions. As shown from the results (Table 2), it was found that the efficiency of vitamin D3 biotransformation into calcitriol, using cell lysate, is much more better than that achieved using intact cells. About 200 fold increase in the amount of calcitriol produced per one million cells per hour was achieved using cell lysate. In addition, a linear relation with, a high rate, between the amount of calcitriol produced and the number of cells was attained upon using sonicated cells (data not shown). The obtained results also give an evidence that the biotransformation process of vitamin D3 , by the test isolate, occurs intracellularly and so it is a function of the diffusion process for both the substrate (vitamin D3 ) and the product (calcitriol). Consequently, the limitation of the biotransformation process, due to diffusion, can be overcome by using cell lysate and this was accomplished in the present study. However, further study is needed in this aspect for evaluation of vitamin D3 biotransformation using crude cell extract, fractionated fraction of crude cell extract as well as characterizing the enzyme(s) involved in the biotransformation process.
Conclusion
From the different conducted experiments, it can be concluded that the test isolate Actinomyces hyovaginalis species A11-2 can transform vitamin D3 into its biologically active form (calcitriol) to give about 180 μg/L calcitriol using the following conditions:
• Preculture medium: Nutrient broth
• Main culture medium consisting of 15 g fructose, 15 g defatted soyabean, 5 g sodium chloride, 2 g calcium carbonate, 1 g dipotassium hydrogen phosphate, 0.2 g sodium fluoride per liter (initial pH of 7.8).
• Substrate (vitamin D3) concentration of 0.2 g/L, added after 2 days of beginning of main culture and remained for 4 days biotransformation time under the growth conditions of 28°C incubation temperature and 200 rpm agitation.
• Replacing deffated soyabean with skim milk can increase calcitriol production by 1.2 fold (220 μg/L).
An alternative promising biotransformation approach for vitamin D3 into calcitriol by the test isolate Actinomyces hyovaginalis A11-2 can be applied depending on the use of cell lysate where about 118 μg calcitriol can be obtained using the lysate of cells obtained from 1 L culture in a time not exceeding 6 hrs.
175
References
- McCollum EVN, Simmonds HT, Parsons PG, Shipley, Park EA (1921) Studies on experimental rickets I: The production of rachitis and similar diseases in the rat by deficient diets. J BiolChem 45: 333 –342.
- Madhok TC, DeLuca HF (1979) Characteristics of the rat liver microsomal enzyme system converting cholecalciferol into 25-hydroxycholecalciferol Evidence for the participation of cytochrome p-450. Biochem J 184: 491-499.
- DeLuca HF, Schnoes HK (1983) Vitamin D: recent advances. Annu Rev Biochem 52: 411–439.
- Ikekawa N (1987) Structures and biological activities of vitamin D metabolites and their analogs. Med Res Rev 7: 333-366.
- Brenza HL, DeLuca HF (2000) Regulation of 25-hydroxyvitamin D3 1α-hydroxylase gene expression by parathyroid hormone and 125-dihydroxyvitamin D3. Arch BiochemBiophys 381: 143- 152.
- Seino YH, Tanaka K, Yamaoka K, Yabuuchi H (1987) Circulating 1α 25-dihydroxyvitamin D levels after a single dose of 1 α 25-dihydroxyvitamin D3 or 1 α -hydroxyvitamin D3 in normal men. Bone Miner 2: 479–485.
- Bouillon R, Okamura H, Norman AW (1995) Structure-function relationships in the vitamin D endocrine System. Endocrine Rev 16: 200-257.
- DeLuca HF (2004) Overview of general physiologic features and functions of vitamin D. Am J ClinNutr 80(6): 1689-1696.
- Larry EJ (2007) “Vitamin D” The Merck manuals https:// wwwmerckmanualscom/professional/sec01/ch004/ch004k [accessed 12/11/2009].
- Grant WB, Holick MF (2005) Benefits and requirements of vitamin D for optimal health. Altern Med Rev 10 (2): 94–111.
- Eastell R, Riggs BL (1997) Vitamin D and osteoporosis In: D Feldman F H Glorieux and J W Pike Vitamin D Academic Press New York pp 695–711.
- Kametani T, Furuyama H (1987) Synthesis of vitamin D3 and related compounds. Med Res Rev 7: 147–171.
- Sasaki J, Mikami A, Mizoue K, Omura S (1991) Transformation of 25- and 1α-hydroxyvitamin D3 to 1α 25-dihydroxyvitamin D3 by using Streptomyces sp strains. Appl Environ Microbiol 57: 2841- 2846.
- Sasaki J, Miyazaki A, Saito M, Adachi T, Mizoue K, Hanada K, Omura S (1992) Transformation of vitamin D3 to lα 25-dihydroxyvitamin D3 via 25-hydroxyvitamin D3 using Amycolatasp strains. ApplMicrobiolBiotechnol 38: 152–157.
- Abass A, Aboshanab KM, Aboulwafa MM, Hassouna NA (2011) Actinomyceshyovaginalis: A novel bacterial isolate with transforming activity of vitamin D3 to 1α 25-dihydroxyvitamin D3. J Am Science. (accepted).
- Miller JH (1972) Experiments in molecular genetics Cold Spring Harbor Laboratory Cold Spring Harbor New York pp: 433.
- Hipolito CN, Crabbe E, Badillo CM, Zarrabal OC, Mora MAM, Flores GP, Cortazar MAH, Ishizaki A (2008) Bioconversion of industrial wastewater from palm oil processing to butanol by Clostridium saccharoper-butylacetonicum N1-4 (ATCC 13564) J Cleaner Prod 16: 632–638.
- Kang DJ, Hong-Sub L, Joon-Tae P, Ji Sun B, Soon-Kwang H, Tae-Yong K (2006) Optimization of culture conditions for the bioconversion of vitamin D3 to 1α 25-dihydroxyvitamin D3 using Pseudonocardiaautotrophica ID 9302. Biotechnol Bioprocess Eng 11(5): 408-413

