Anahtar Kelimeler
Ekosistem, Hamsi, Karadeniz, Ötrofikasyon, Mnemiopsis, Beroe
Keywords
Anchovy, Black Sea, Ecosystem, Eutrophication, Mnemiopsis, Beroe
Giri?
Türkiye 1981'den itibaren tüm Akdeniz ve Karadeniz ülkeleri aras?nda, her y?l en fazla bal?k avlayan ülke olmu?tur. Karadeniz'in Türk bal?kç?l???nda özel bir yeri vard?r. 1970'den beri Türkiye'nin y?llara göre yakalad??? toplam bal?k miktar?n?n %70'den fazlas? bu denizden elde edilmi? olup, Karadeniz'den yakalanan hamsi (Engraulis encrasicolus), av?n büyük bir k?sm?n? (1980 ve 1988 y?llar? aras?nda toplam av?n % 60-72'sini) olu?turmu?tur. 1975'lerden sonra hamsi ve zooplanktonla beslenen di?er pelajik bal?k populasyonlar? sürekli olarak artm??t?r. Ancak 1988'den sonra hamsideki ve dolay?s?yla Türk bal?kç?l???ndaki ani bir dü?ü? hemen dikkati çekmektedir. Resmi istatistiklere göre (Anonim, 1968-1990) Türkiye'nin Karadeniz'de avlad??? hamsi miktar? 1988'deki 295 bin ton de?erinden 1989'da 97 bin tona ve 1990'da 66 bin tona dü?mü?tür. Bu, 1988 ve 1990 y?llar? aras?nda yakla??k 4 kat azalma demektir.
Son y?llarda Karadeniz’deki ekolojik denge de?i?imi sonucunda birçok tür bu ortamda hakimiyet sa?lamaya ba?lam??t?r. Karadeniz 1960’l? y?llardan bu yana çe?itli çevresel problemler ile kar??la?m??t?r (Zaitsev, 1992). A??r? avlanma, ötrofikasyon, oksijen yetersizli?i gibi nedenlerden dolay? son y?llarda baz? zooplankton türleri ya ortadan kaybolmakta ya da nadiren bulunmaktad?r (Zaitsev ve Alexandrov, 1995; Caddy, 1993; Gerlach, 1990). Buna kar??l?k di?er baz? organizmalar geli?mekte ve büyük miktarlara ula?maktad?r. Örne?in 1980’lerin ba??nda Karadeniz’e do?u ABD’den kargo gemilerinin balans suyunda ta??nan tarakl? Mnemiopsis leidyi, Karadeniz’de bütün ekosistemi etkilemi?tir (Kideys, 1994; K?deys, 2002). 1989 y?l?n?n yaz ay?nda doymak bilmez bu zooplanktonik predatör çok geni? biyokütle de?erine (>1kg m2) ula?m??t?r (Vinagradov ve ark., 1989). Mnemiopsis herbivor zooplankton üzerinden oburca beslenerek ve yemesiyle orant?l? büyüme ve üreme özellikleri ile Karadeniz pelajik ekosistemini önemli ölçüde etkilemi?tir. Bu durum indirekt olarak bütün Karadeniz çana??ndaki birinci üretimin ve fitoplankton biyokütlesinin artmas?na neden olmu?tur. Mnemiopsis Karadeniz’in bask?n bal??? hamsi Engraulis encrasicolus türünün yem zooplankton için rekabete girmesi ve hamsi yumurtalar?n? ve larvalar?n? tüketmesi Karadeniz’de hamsi ve di?er pelajik bal?klar?n aniden azalmas? için önemli nedenlerden birisi olmu?tur (Kideys, 1994; Kideys ve ark., 2000). 1997’de görülen ve muhtemelen yine kuzeybat? Atlantik kaynakl? oldu?una inan?lan Beroe ovata adl? ba?ka bir ktenofor türü, Mnemiopsis üzerinden beslenerek Mnemiopsis türünün Karadeniz’deki hakimiyetine son verirken, ekosistemin eski haline dönmesine çok büyük katk? sa?lam??t?r (Finenko ve ark., 2001). Mnemiopsis populasyonun azalmas? jelatinsiz zooplankton, hamsinin avc?l??? (Kideys ve ark., 2000) ve yumurta yo?unlu?unun (Kideys ve ark., 1999) artmas?na neden olmu?tur.
Son yirmi y?ld?r çok h?zl? de?i?ikliklere maruz kalan Karadeniz’de önemli miktarda çal??ma olmas?na ra?men, bu bölgede fitoplankton, zooplankton, ihtiyoplankton ve makrozooplanktonun mevsimsel da??l?m?n? tüm y?l boyunca düzenli aral?klarla veren bir çal??ma yoktur. Bu nedenle kalitatif ve kantitatif çal??malar?n sürekli olarak yap?lmas?, biyolojik çe?itlili?in korunmas?, anla??lmas? ve meydana gelebilecek de?i?ikliklerin izlenmesi için zorunludur.
Karadeniz’in genel özellikleri
Fiziksel o?inografi ve derinlik
Karadeniz; 40°- 46° N enlemleri ile 27°- 41° E boylamlar? aras?nda bulunan yar? kapal? bir iç denizdir. Güneybat?da Türk Bo?azlar Sisteminin olanak verdi?i miktarda dünya denizleriyle ba?lant?s? bulunur. Bu k?s?tl? su de?i?imi, sadece yüzeyden 150 m derinli?e kadar (toplam hacmin % 15’i) oksijen içeren, daha derinde ise hidrojen sülfür bulunduran neredeyse tamam? oksijensiz bir ortam?n olu?mas?na yol açar. Yüzey alan? 4.2 x 105 km2 ve hacmi 5.3 x 105 km3 olan Karadeniz, dünyan?n en büyük anoksik basenidir. Karadeniz’in derin düz taban? (<2000 m) toplam alan?n % 60’?ndan fazlas?n? kapsar. En derin yeri yakla??k 2300 m olup ortalama derinli?i 1240 m olarak hesaplanm??t?r (Ross ve ark., 1974).
K?y? topografyas? k?y?ya paralel, yakla??k 20 km enine bir ku?ak boyunca oldukça belirgin de?i?imler gösterir. Dinyeper, Dinyester ve Tuna gibi büyük nehirlerin denize döküldü?ü Kuzeybat? Karadeniz bölgesinde geni? bir k?ta sahanl??? vard?r. Geni? kuzeybat? sahanl??? yakla??k 100 m derinli?e kadar K?r?m yar?madas? ile Karadeniz’in bat? k?y?s? aras?nda yer al?r ve bat?- güneybat? k?y?lar? boyunca güneye uzan?r. Süreklilik gösteren bu düz k?ta sahanl???n?n eni güneye do?ru azal?r ve derinli?in 100 m den birden 1500 m ye indi?i Sakarya Kanyonu’nda sonlan?r. Güney k?y?lar?nda Sakarya, Ye?il?rmak ve K?z?l?rmak nehirlerinin bo?ald??? bölgelerde küçük ölçekli yöresel k?ta sahanl?klar? bulunur (Sorokin, 1983; Ünlüata ve ark., 1990; O?uz ve ark., 1993).
Su kütlesi, dikey tabakala?ma ve su sirkülasyonu
Karalarla çevrili büyük, kapal? bir basen olu?u nedeni ile Karadeniz’de toplam su kütlesinin bütçesi ve hidrokimyasal yap?s? kritik olarak hidrolojik dengenin elementlerine ba?l?d?r. Yüzey sular?n?n karakteristi?i, temelde tatl? su girdisi taraf?ndan kontrol edilmektedir ve s?? ?stanbul bo?az? boyunca gerçekle?en al?? veri? oldukça k?s?tl?d?r. Di?er yandan, daha derin sular?n havalanmas? ve haloklinin yap?s? ise yine ?stanbul Bo?az?ndan giren Akdeniz sular? ile yak?ndan ili?kilidir.
Büyük bir k?sm? Tuna, Dinyeper ve Dinyester nehirlerinden sa?lanan tatl? su girdisi (~ 400 km3 y?l-1) buharla?ma yolu ile gerçekle?en su kayb?ndan (350 km3 y?l-1) daha yüksek oldu?u için Karadeniz pozitif bir su dengesine sahiptir. Rapor edilen tahminler büyük de?i?iklikler göstermesine ra?men literatür taramas?na dayanarak su dengesi yakla??k: Ya??? için ~350 km3 y?l-1, tatl? su girdisi için ~ 400 km3 y?l-1 ve yüzeyden buharla?ma için ~350 km3 y?l-1 olarak bildirilmi?tir. Su bütçesindeki geri kalan bile?enin ise ?stanbul Bo?az?ndan gerçekle?en net ak?nt?ya kar??l?k geldi?i bildirilmi?tir (Tolmazin, 1985; Ünlüata ve ark., 1990; Bingel ve ark., 1993). Kuzeybat? sahanl???ndaki üç büyük nehir, Karadeniz'de y?ll?k 400 km3 olarak tesbit edilen toplam nehir girdisinin %67'sini sa?lar (Tuna tek ba??na yakla??k 203 km3 y?l-1, Dinyeper 54 km3 y?l-1 ve Dinyester 9,3 km3 y?l-1). Bu üç nehrin y?ll?k ak?nt?s?, s?? sahanl?k hacminin %4'üne yakla??r. Birçok ülkenin zirai ve sanayi alanlar?n?n besledi?i çok büyük bir drenaj havzas?, bahsedilen nehirlerin ak?nt?s?na katk?da bulunur.
So?uk ve az tuzlu sular?n daha s?cak ve tuzlu sular?n üzerinde yer ald??? Karadeniz özgün bir tabakala?ma yap?s? gösterir. Yüzeydeki az tuzlu sular tatl? su girdisinin fazla olmas? ile olu?mu?tur, derinlerdeki tuzlu sular ise Akdeniz etkisini yans?t?rlar. Tuzluluk tabakala?mas?n?n konveksiyonu s?n?rlay?c? etkisi nedeniyle tuzluluk ara yüzeyi ‘haloklin’ ile yo?unluk ara yüzeyi ‘piknoklin’ yakla??k 100-200 m derinlik aral???nda yer al?r ve 8°C s?n?r izotermi ile belirlenen So?uk orta sular?n (Cold Intermediata Water, CIW) alt s?n?r? ile çak???rlar. Benzer mekanizmalarla gerçekle?en çe?itli özelliklerin dikey de?i?imi sonucunda, oksijen ara yüzeyi ‘oksiklin’ ve kimyasal ara yüzey ‘kemoklin’ de ayn? derinliklerde yer al?r. Kimyasal tabakala?man?n, suboksik bölge (oksik ve anoksik bölgeler aras?nda bir geçi?) ve partikül tabakalar? vs. gibi, daha ince ayr?nt?lar? bulunmaktad?r (Murray ve ark., 1993). Tuzlulu?u dü?ük (~ ‰ 18) ince bir kar???m tabakas? (~30 m) yüzeyde çok güçlü bir mevsimsel ?s?nma ve so?umaya u?rar. Minimum çekirdek s?cakl??? ~6°C olan so?uk ara sular taraf?ndan (CIW) karakterize edilen, so?uk orta tabaka (CIL), sabit haloklin ile mevsimsel termoklin aras?nda yer al?r. So?uk ara sular yaz?n ?l?k bir yüzey tabakas? ile kapland???ndan so?uk ara tabaka, yüzey alt? s?cakl?k minimumu ?eklinde ortaya ç?kar. K???n Karadeniz’in hemen hemen her taraf?nda ve Bat? Karadeniz’in Anadolu k?y?lar?nda so?uma ve konveksiyon nedeni ile 70-80 m derinlere kadar inebilen ve minimum s?cakl??? 6-7°C olabilen izotermal bir tabaka meydana gelir. S?cakl?k ve tuzluluktaki mevsimsel ve y?ll?k de?i?kenlik 500 m derinli?e kadar devam eder. 500 m alt?ndaki derin sular? temelde dura?and?r (Özsoy ve ark., 1986 ve 1993). Yerel karars?zl?klar?n ince yap?lar olu?turabildi?i s?n?rlara yak?n bölgeler d???nda, özelliklerde çok büyük de?i?meler gözlenmez (Özsoy ve Be?iktepe, 1995). 1700 m’lik bir derinli?in alt?nda deniz taban?ndan kaynaklanan jeotermal ?s?nman?n sürücü gücü ile olu?an ve ~ 400 m kal?nl???nda olan bir taban konveksiyon tabakas? mevcuttur (Özsoy ve ark., 1993; Murray ve ark., 1993, Özsoy ve Be?iktepe, 1995).
Geçmi?te yap?lan ve günümüzde uydu analizleri ile detayl? o?inografik çal??malar?n sonucunda desteklenip geli?tirilen incelemeler neticesinde, siklonik bir s?n?r ak?nt?s? (S?rt Ak?nt?s? (Rim Current): O?uz ve ark., 1992 ve 1993’da s?n?r ak?nt?s?na verilen isim) Karadeniz’in genelindeki dola??m?n ana özelli?idir (O?uz ve ark., 1996). Bu temel dola??m sistemi; merkezi k?sm? i?gal eden, siklonik, iki veya üç hücresi bulunan, bunlar?n çevresi boyunca bir dizi antisiklonik girdaplar? ve basenin en do?u kö?esinde yer alan bir antisiklonik dola??m? (Batum Girdab?) içeren karma??k bir sistemdir. Bunun yan?nda ?stanbul Bo?az?n?n bat?s?nda, Sinop’un do?usunda ve K?r?m Yar?madas?n? her iki taraf?nda, S?rt Ak?nt?s?n?n k?y?lar?nda bulunan çe?itli antisiklonik girdaplar gözlenmi?tir (O?uz ve ark., 1993) (?ekil. 1).
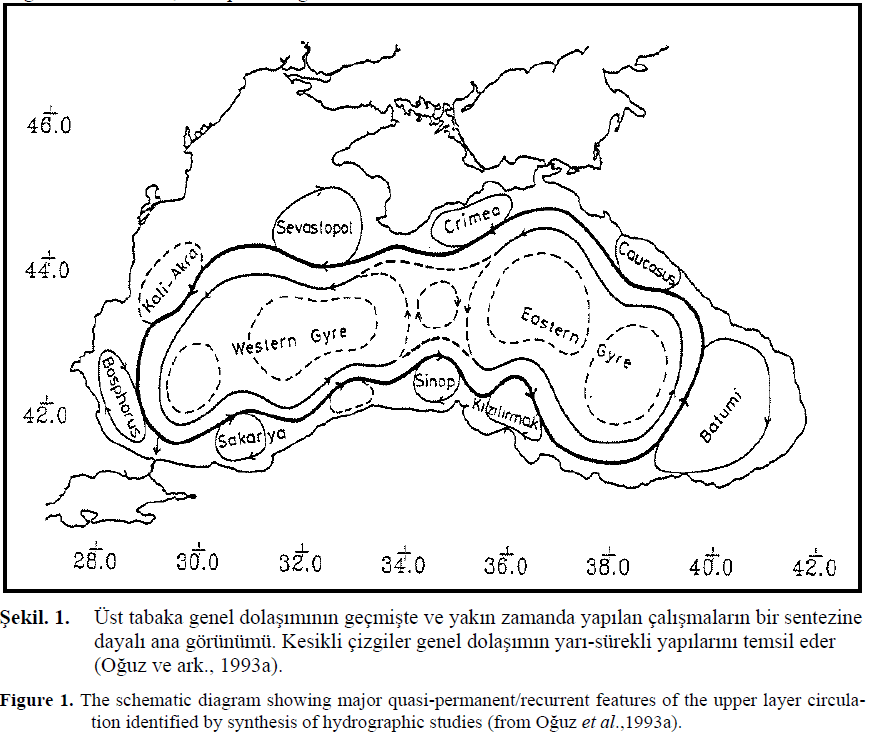
?ekil. 1. Üst tabaka genel dola??m?n?n geçmi?te ve yak?n zamanda yap?lan çal??malar?n bir sentezine dayal? ana görünümü. Kesikli çizgiler genel dola??m?n yar?-sürekli yap?lar?n? temsil eder (O?uz ve ark., 1993a).
Figure 1. The schematic diagram showing major quasi-permanent/recurrent features of the upper layer circulation identified by synthesis of hydrographic studies (from O?uz et al.,1993a).
Besin tuzlar? girdisindeki de?i?imler
Son çeyrek yüzy?lda, Karadeniz'in bilhassa kuzey ve bat? kesimlerindeki besin tuzu yo?unluklar? önemli miktarlarda artm??t?r. Karadeniz'e besin tuzlar?n?n ta??nmas?nda, atmosfer önemli bir kaynak olmakla beraber, nehirlerin rolü büyüktür. Maalesef atmosfer yoluyla bu denize ta??nan besin tuzu miktarlar? bilinmemektedir, ancak belli ba?l? nehirler yoluyla ta??nan y?ll?k girdi miktar? hakk?nda birtak?m çal??malar mevcuttur.
417000 km3 lük sulama alan?yla Tuna nehri Karadeniz için en önemli gübre kayna??d?r. Tuna yoluyla Karadeniz'e ta??nan (fosfat) fosfor miktar? 1950'lerdeki y?lda 12.6 tondan 1987'de 30.4 tona yükselmi?tir (Bodeanu, 1989). Y?ll?k nitrojen miktar? daha da fazla bir art?? göstermi?tir (1950'lerde 143000 ton, 1988'de 741000 ton). Fosfor ve nitrojen artarken silikonun y?ll?k girdisi bir azalma e?ilimi gösterir. Bu durum, nehrin veya onun kollar? üzerinde yap?lm?? olan çok say?da barajlardan kaynaklanmaktad?r (Tablo 1).
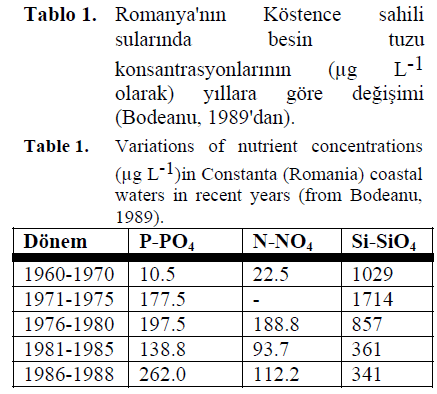
Tablo 1. Romanya'n?n Köstence sahili sular?nda besin tuzu konsantrasyonlar?n?n (μg L-1 olarak) y?llara göre de?i?imi (Bodeanu, 1989'dan).
Table 1. Variations of nutrient concentrations (μg L-1)in Constanta (Romania) coastal waters in recent years (from Bodeanu, 1989).
Benzeri e?ilimler di?er nehirlerin besin tuzu yükünde de gözlenmi?tir. Örne?in, 1952 ve 1977 y?llar? aras?nda, Dinyester nehrinin denize yak?n k?s?mlar?nda, nitrit, nitrat, fosfat ve silikon konsantrasyonlar? (litrede mikrogram olarak), s?ras?yla, 0-20'den 36-150'ye, 0- 1000'den 400-3000'e, 0-50'den 15-260'a ve 1000-5600'den 2300-9200'e yükseltmi?tir (Tolmazin, 1985).
Son çeyrek yüzy?l boyunca, Karadeniz'in aç?k sular?nda maksimum nitrat konsantrasyonlar? 1960'lardaki 2-4 μM'dan (0.1-0.3 μg l-1) yava? yava? 1978-1980'de 5-7 μM'a (0.4-0.5 μg l-1) ve son olarak 1988- 1991'de 8-9 μM'a (0.6-0.7 μg l-1) yükselmi?tir (Codispoti ve ark., 1991).
Fitoplankton
Besin tuzu dengesindeki en ufak bir de?i?iklik bile önce fitoplankton komünitelerinde ve daha sonra da kar???k besin a?? olay? nedeniyle tüm ekosistemde de?i?imlere neden olabilir.
Karadenizde plankton komünitelerinin ötrofikasyona tepkisinin, artan red-tide ve patlamalar?n da içerildi?i kalitatif ve kantitatif yap?lar?ndak? de?i?imler olarak yans?t?ld??? rapor edilmi?tir. Örne?in, son çeyrek yüzy?lda diatomelerin dinoflagellatlara oran? birçok bölgede de?i?mi?tir. Bologa (1986), diatomlar?n oran?n?n 1960-1970'deki % 67'den (209 tür) 1972-77 peryodunda %46'ya (172 tür) dü?mesi nedeniyle, fitoplankton gruplar?nda kalitatif bir de?i?ime dikkat çekmi?tir (Tablo 2). Ayn? süreç içerisinde dinoflagellat türlerinin say?s? 60 dan 77' ye ç?km??t?r. Fitoplanktondaki bu tip kalitatif de?i?iklikler, ekosistemde yeni türlerin ortaya ç?kt???n? gösterir. Ortamda ilk defa beliren bu türler aras?nda Gonyaulax polygramma (Dinophyceae), Raciborshiella salina (Volvocales) ve Eutreptia lanowii (Eugleninae) yak?n zamanlarda Karadeniz'de yüksek yo?unluklarda rapor edilmi?tir (Mihnea, 1985). Bir diatom olan Hemiaulus hauckii Karadeniz'in güneydo?u k?y?lar?nda a??r? oranlarda gözlenmi?tir (Feyzio?lu, 1990). Bu türün oligotrofikten ötrofik özelli?e geçen sular aras?nda bir geçi? türü oldu?u bildirilmi?tir (Kimor, 1985). Kantitatif olarak, Romanya fitoplank-tonunda diatomelerin oran? 1960- 70'deki % 92.3'ten 1983-1988'de %62.2'ye dü?erken, dinoflagellatlarin oran? ayn? periyot içerisinde 7.6'dan 30.9'a ç?km??t?r (Tablo 3).

Tablo 2. Romanya sahillerinde fitoplankton gruplar?n?n taksonomik da??l?m? (Bologa ,1986; *Bat ve ark., 2005’ten).
Table 2. Taxonomic distrubition of phytoplankton groups in Russian coast (Bologa ,1986 in *Bat et al., 2005).
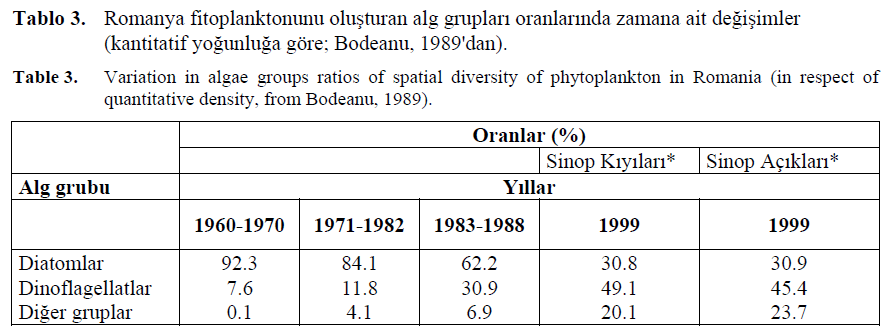
Tablo 3. Romanya fitoplanktonunu olu?turan alg gruplar? oranlar?nda zamana ait de?i?imler (kantitatif yo?unlu?a göre; Bodeanu, 1989'dan).
Table 3. Variation in algae groups ratios of spatial diversity of phytoplankton in Romania (in respect of quantitative density, from Bodeanu, 1989).
Türko?lu (1998) 1995-1996 periyodunda, Sinop Yar?madas? littoral sular?nda toplam 197 taksona ait 1 Cyanophyceae, 83 Dinophyceae, 1 Prymnesiophyceae, 5 Dictyochophyceae, 88 Bacillariophyceae, 1 Euglenophyceae, 1 Acantharea ve 17 Ciliata türü tespit etmi?tir.
Ötrofikasyonun bir di?er kaç?n?lmaz sonucu fitoplanktonun hücre say?s?nda ve biyomas?ndaki art??t?r. Örne?in Mihnea (1985), diatom Skeletonema costatum'un litredeki hücre say?s?n?n, 1962-1965'teki 1 x 104 - 4 x 106'den 1984 te 8.3 x 107'ye yükseldi?ini rapor etmi?tir. Benzer ?ekilde dinoflagellat Exuviella (=Prorocentrum) cordatum 1960'larda birkaç milyon iken, 1975-1983'de 1 x 107 - 1 x 108 hücre L-1 de?erine yükselmi?tir. Karadenizin Romanya sahillerinde son 20-30 yilda yap?lan çal??malar çok say?da fitoplankton a??r? üremelerini rapor etmektedir. Örne?in, fitoplanktonun 1983-1988'deki ortalama biyomas?, 1959-1963' tekinden 10 kat daha fazlad?r. 1960-1970' de yüksek say?sal yo?unluklara eri?ebilen (1 x 104 hücre L-1) tür say?s? sadece 38 iken, 1971-1982'de 61'e ve 1983-1988'de 72'ye yükselmi?tir. Yak?n geçmi?te tüm denizde Exuviaella cordata redtide'lar? ço?alm??t?r. 1986'daki a??r? üreme s?ras?nda, Burgaz Körfezi’ndeki Exuviaella biyomas? 1 x 109 hücre L-1 (1 g L-1) ye ula?m??t?r (Sukhanova ve ark., 1988). Red-tide yapan di?er bir dinoflagellat olan Noctiluca scintillans Tuna nehri a?z?nda 100 g L-1 gibi büyük bir yo?unlu?a ula?m??t?r ( Vinogradov ve ark., 1989'dan). 1976-1977' den beri zaman zaman Noctiluca a??r? ço?almalar? belli bölgelerin ola?an bir özelli?i haline gelmi?tir (Caddy ve Griffiths, 1990). Fitoplankton patlamalar? dikey ???k geçirgenli?ini etkiler ve çözünmü? oksijen içeri?inde azalmaya neden olur ki bu Karadeniz’de oksijensiz tabakan?n üst s?n?r?nda yükselmeye götürebilir.
Zengin besin tuzu girdilerinin etkisinde bulunan Karadeniz'in orta derecede (zaman ve mekana ba?l? olarak bazen de yüksek derecede) verimlili?e sahip oldu?u bilinmektedir (Koblentz-Mishke ve ark., 1970). Sorokin (1983)' e göre bu denizde iki fitoplankton patlamas? (=bloom) dönemi bulunmaktad?r. Bunlardan birincisi erken ilkbaharda diatomlar taraf?ndan, ikincisi ise sonbaharda kokkolitler taraf?ndan gerçekle?tirilmektedir. Bu nedenle bu çal??mada elde edilen dü?ük hücre yo?unlu?u de?erleri normaldir. Bunun yan?nda sahil bölgelerinde kokkolit ya da dinoflagellatlar taraf?ndan olu?turulan küçük çapl? patlamalar da bilinmektedir. Yine Kuzeybat? k?ta sahanl??? bölgesinde klasik kurallara uymayan beklenmedik yaz patlamalar? olu?maktad?r (Bologa, 1986; Benli, 1987). Bu patlamalardan baz?lar? "red tide" halinde olmaktad?r (Sorokin, 1983; Sukhanova ve ark. 1988). K?? aylar?na ait s?n?rl? veri olmakla birlikte Karadeniz'in Bat? Anadolu k?y?lar? boyunca zaman zaman büyük çapl? ço?almalar?n oldu?u belirlenmi?tir (Sur ve ark. 1994; Uysal ve Sur, 1995).
Fitoplankton kommunitelerinde meydana gelen patlamalarda diatomelerin rolünün azalmas?, bununla birlikte dinoflagellat, euglenoid ve kokkolitoforidlerin rolünün artmas? Karadeniz’e bo?alan nehirlerin etkisi sonucu son çeyrek yüzy?lda antropojenik girdilerle birlikte nütrientlerin de birikmesinden kaynakland??? tahmin edilmektedir. Önceki çal??malardan elde edilen sonuçlarda hem kalitatif hem kantitatif bak?mdan diatomeler üstün iken, ?imdiki çal??malar dinoflagellatlar?n üstün durumda oldu?unu göstermektedir. Örne?in Leptocyclindrus danicus, Ditylum brightwellii ve di?er büyük hacimli türlerin olu?turduklar? populasyonlar gerileme gösterirken Prorocentrum micans ve Emiliania huxleyi gibi küçük hacimli türlerin olu?turduklar? populasyonlar büyük kütlesel geli?im gösterirler (Zaitsev ve Aleksandrov, 1997). Sinop yar?madas? littoral sular?nda yapt???m?z bu ara?t?rmada Exuviaella cordata, Exuviaella compressa, Exuviaella marina, Prorocentrum micans, Scrippsiella trochoidea, Heterocapsa triquetra, Gymnodinium sp., Gymnodinium splendens, Thalassiosira sp., Nitzschia delicatissima, Nitzschia tenuirostris, Hillea fusiformis, Glenodinium paululum, Emiliania huxleyi, Anabaena sp. ve küçük kamç?l?lar türleri her örnekleme ay?nda yo?un bir ?ekilde bulunmu?lard?r. Bu da Yaz aylar?nda a??r? ço?alan türlerin K?? dönemlerinde de a??r? ço?alabildiklerini göstermektedir.
Morozova ve Wodjanitkaja (1963, Eker ve ark., 1999’dan), yapt?klar? çal??mada Karadeniz’de 350 fitoplankton türü tespit etmi?ler ve toplam tür say?s?n?n % 42’sini diatomlar?n, % 41’ini se dinoflagellatlar?n olu?turdu?unu bildirmi?lerdir.
Petrova-Karadjova (1973), yapt?klar? çal??mada Bat? Karadeniz’de 255 tür tespit etmi?, bu türlerin % 42’sinin diatom, % 39.6’s?n?n dinoflagellat oldu?unu rapor etmi?lerdir.
Uysal (1993), 1989 y?ln?n?n Nisan ay?nda Güney Karadenizde toplam 87 istasyonda 92 fitoplankton türü tespit etmi?, toplam tür say?s?n?n % 60’?n? diatomlar?n, % 20’sini dinoflagellatlar?n olu?turdu?unu belirtmi?tir. Bunun yan?nda ?ubat 1990’da yapt??? örneklemede toplam 148 istasyonda 122 tür tespit etmi? bu örnekleme peryodunda da diatomlar?n dinoflagellatlardan tür say?s? bak?m?ndan çok oldu?unu belirtmi?tir. Nisan 1990’daki örneklemesinde ise tespit etti?i 56 fitoplanktonun % 52’sini diatomlar?n, % 30’unu da dinoflagellatlar?n olu?turdu?unu rapor etmi?tir.
Bayraktar (1994), Do?u Karadeniz Bölgesi’nde yapt??? çal??mas?nda toplam 136 istasyonda 118 fitoplankton türü tespit etmi?, bu türlerin 76 s?n?n dinoflagellat ve 38’inin diatom oldu?unu bildirmi?tir.
Karaçam ve Düzgüne? (1990), Kas?m 1987- Ekim 1988 tarihleri aras?nda yapt?klar? ara?t?rmada yüzey deniz suyunda toplam 17 diatom ve 12 dinoflagellat türü tespit etmi?lerdir.
Feyzioglu (1990), Temmuz 1989 döneminde yapt??? çal??mada 38 diatom ve 26 dinoflagellat türü rapor etmi?tir. Yine, Feyzio?lu (1996), çal??mas?nda 102 fitoplankton türü tespit etmi? bunlar?n 56’s?n?n diatom, 35’inin dinoflagellat oldu?unu belirtmi?tir.
Uysal ve ark. (1997), yapt?klar? ara?t?rmada Nisan 1996 döneminde 73, Temmuz 1996 döneminde 119 fitoplankton türü tespit etmi?ler ve toplam tür say?s?n?n Nisan dönemi içinde % 45’ini, Temmuz dönemi içinde ise % 60’?n? dinoflagellatlar?n olu?turdu?unu rapor etmi?tir.
Eker ve ark. (1999), yapt?klar? ara?t?rmada toplam 122 istasyonda örnekleme peryodu (Mart-Nisan 1995 ve Ekim 1995) boyunca 142 fitoplankton türü tespit etmi?ler ve bu türlerin 59’unun dinoflagellat, 41’inin diatom oldu?unu rapor etmi?lerdir.
Türko?lu ve Koray (2002), A?ustos-1995 ve Temmuz-1996 dönemleri aras?nda Güney Karadeniz k?y?lar?nda yapt?klar? çal??mada toplam 178 tür tespit etmi?lerdir. Bu türlerin 88’inin diatomlara, 83’ünün dinoflagellatlara ait oldu?unu rapor etmi?lerdir.
Di?er çal??malarla kar??la?t?r?ld??? zaman Bat ve ark’n?n bulduklar? tür say?s? (2002 y?l?nda 107 tür, 2003 y?l?nda 94 tür) yaln?zca iki istasyonda çal???lmas?na ra?men oldukça yüksek bir say?d?r. Yap?lan di?er çal??malara bak?ld???nda örnekleme istasyonunun çoklu?u dikkati çekmektedir. Güney Karadeniz’in Sinop Burnu bölgesinde tür kompozisyonu bak?m?ndan yap?lan çal??malar oldukça azd?r (Tablo 4.)
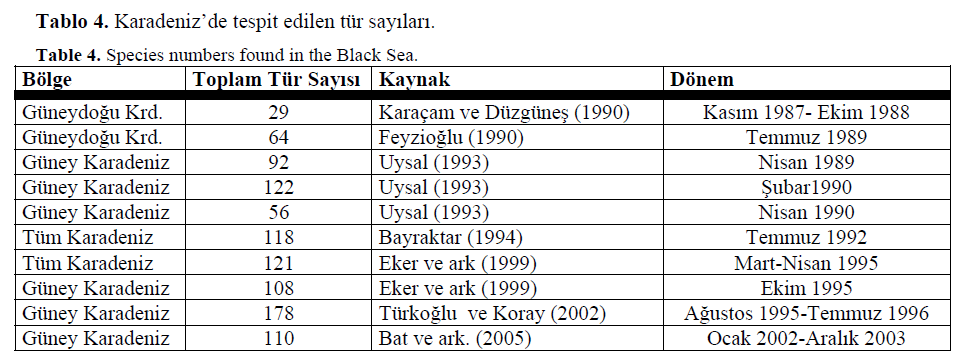
Tablo 4. Karadeniz’de tespit edilen tür say?lar?.
Table 4. Species numbers found in the Black Sea.
Dünya denizlerinde ve okyanuslar?nda ve hatta Kuzey Karadeniz’e k?y?s? olan ülkelerde uzun y?llardan beri fitoplanktonik organizmalar?n yap?lar?ndaki kalitatif ve kantitatif de?i?imler sürekli takip edilmektedir. Buna kar??l?k fitoplanktonun Karadeniz’in ülkemiz k?y?lar?ndaki durumu ve de?i?imine verilen önem ancak son y?llarda artm??t?r.
Fitoplanktonik organizmalar küçük olmalar? ve ortam ?artlar?na h?zl? tepki göstermeleri nedeniyle fiziksel veya kimyasal o?inografide meydana gelebilecek küçük de?i?imlerden etkilenmesi kaç?n?lmazd?r. De?i?en ortam ?artlar? fitoplanktonik yap? içerisinde meydana gelen nitel ve nicel farkl?l?klar ile kar??l?k bulacakt?r. Bu sebeple belli bir bölgede bulunan fitoplanktonik yap? içerisindeki indikatör türlerin tespit edilmesi ile ekosistemde meydana gelen de?i?imlerin anla??lmas?nda yard?mc? olacakt?r. Yap?lan çal??malar süreklilik ta??rsa yap?lacak yorumlar do?ru ve etkili olur.
Zooplankton
Do?al olarak besin zincirinde ya da a??nda yer alan organizmalardan herhangi birinin yok edilmesi ekosistemdeki dengeyi olumsuz yönde etkileyecektir. Örne?in Karadeniz’de ya?ayan tüm canl?lar ve bunlar?n aras?ndaki ili?kiler yuma?? diye tan?mlayabilece?imiz besin a??nda; önemli bir bal?k türü olan hamsiyi ele alacak olursak; 1980’li y?llar?n sonunda ve 1990’l? y?llar?n ba??nda a??r? art?? gösteren ve doymak bilmez bir zooplankton avc?s? olan ktenofor Mnemiopsis sp., hamsinin yumurta ve larvalar? üzerinden beslenir. Bununla birlikte Mnemiopsis leidyi hamsinin severek tüketti?i Oithona nana ve Paracalanus parvus gibi kopepod türlerini önemli ölçüde tüketerek hamsi ile rekabete girmi?tir. Bu küçük kopepodlar?n yok olmas?ndan dolay? bal?k larvalar? biraz daha büyük kopepod türlerini tüketmeye ba?lam??t?r (Tkach ve ark., 1998). Kideys ve ark. (2000), 10 mm uzunlu?undaki Engraulis encrasicolus larvas?n?n ba??rsa??nda 0.75 mm uzunlu?unda kopepod Acartia clausi türünü tespit etmi?lerdir.
Besin de?eri aç?s?ndan da planktonik organizmalar büyük önem ta??maktad?rlar. Larvalar?n ilk geli?im evrelerinde, boyutlar?na uygun O. nana ve P. parvus tüketemeyi?leri sonucunda, özellikle II. ve III. evredeki larvalar?n hayatta kalabilme ?ans?n?n azald??? tahmin edilmektedir (Kideys ve ark., 2000).
Ekonomik de?eri fazla olan bal?klar?n hemen hepsi pelajik olarak yumurtadan ç?kmakta ve ya?amlar?na plankton olarak ba?lamaktad?r. Besin keselerini tüketen bal?k larvalar?n?n hayatta kalabilmeleri, plankton populasyonuna ba?l?d?r (Özel, 1998). Bu durumda ortamda meydana gelebilecek herhangi bir de?i?iklik planktonu etkiledi?i gibi bal?k populasyonunu da do?rudan etkileyecektir. Bu nedenle bal?klar için besin olu?turan planktonun en zengin oldu?u bölgeler zengin bal?kç?l?k alanlar?n? olu?turmaktad?r. Plankton çal??malar?n?n bal?k stoklar?nda optimum avlanma veriminin belirlenmesinde önemli bir yeri vard?r. Bir bölgedeki toplam bal?k stoklar?n?n ara?t?r?lmas?, yumurta ve larvalardaki ölüm miktar?n?n saptanmas? ile mümkün oldu?u gibi ayn? zamanda da planktonik canl?lara ba?l?d?r (Gürtürk, 1962). Organik madde ve buna ba?l? olarak planktonca zengin olan Karadeniz’den, Türkiye’deki bal?k üretiminin %82’sinin sa?lanmas? bunun bir göstergesi say?labilir (Anonim, 1998).
Sucul ortamda bulunan canl?lar do?al bir dengede varl?klar?n? sürdürürler. Son y?llarda Karadeniz ekolojik denge de?i?ime u?ram??t?r. Bu de?i?im sonucunda birçok tür bu ortamda hakimiyet sa?lamaya ba?lam??t?r. A??r? avlanma, ötrofikasyon, oksijen yetersizli?i gibi nedenlerden dolay? son y?llarda baz? zooplankton türleri ortadan kaybolmakta veya az bulunmaktad?r (Gerlach, 1990; Caddy, 1993; Zaitsev ve Alexandrov, 1995). Buna kar??l?k di?er baz? organizmalar geli?mekte ve büyük miktarlara ula?maktad?r. Örne?in 1980’lerin ba??nda Karadeniz’e do?u ABD’den kargo gemilerinin balast suyunda ta??nan tarakl? deniz anas? Mnemiopsis, Karadeniz’de bütün ekosistemi etkilemi?tir (Kideys, 1994; Kideys, 2002). Doymak bilmez bu zooplanktonik predatör 1989 y?l?n?n yaz ay?nda çok geni? biyokütle de?erine (>1kg/m2) ula?m??t?r (Anonim, 1998). Mnemiopsis herbivor zooplankton üzerinden oburca beslenerek ve yemesiyle orant?l? büyüme ve üreme özellikleri ile Karadeniz pelajik ekosistemini önemli ölçüde etkilemi?tir. Bu durum dolayl? olarak bütün Karadeniz çana??ndaki birincil üretimin ve fitoplankton biyokütlesinin artmas?na neden olmu?tur (Kideys ve Romanova, 2001). Mnemiopsis türü Karadeniz’in bask?n bal??? hamsi (Engraulis encrasicolus) ile besin zooplankton için rekabete girmesi, hamsi yumurta ve larvalar?n? tüketmesinden dolay? Karadeniz’de hamsi ve di?er pelajik bal?klar?n aniden azalmas? için önemli nedenlerden birisi olmu?tur (Kideys, 1994; Kideys ve ark., 2000). Ekim 1997’de görülen ve muhtemelen kuzeybat? Atlantik kaynakl? oldu?una inan?lan Beroe adl? ba?ka bir ktenofor türü, Mnemiopsis üzerinden beslenerek Mnemiopsis türünün Karadeniz’deki hakimiyetine son verirken, ekosistemin eski haline dönmesine çok büyük katk? sa?lam??t?r (Finenko ve ark., 2001). Mnemiopsis populasyonunun azalmas? jelatinsiz zooplankton, hamsi avc?l??? (Kideys ve ark., 2000) ve yumurta yo?unlu?unun (Kideys ve ark., 1999) artmas?na neden olmu?tur.
Son yirmi y?ld?r çok h?zl? de?i?ikliklere maruz kalan Karadeniz’de önemli miktarda çal??ma olmas?na ra?men, bu bölgede zooplanktonun mevsimsel da??l?m?n? tüm y?l boyunca düzenli aral?klarla veren bir çal??ma yok denecek kadar azd?r. Bu nedenle kalitatif ve kantitatif çal??malar?n sürekli olarak yap?lmas?, biyolojik çe?itlili?in korunmas?, anla??lmas? ve meydana gelebilecek de?i?ikliklerin izlenmesi için zorunludur.
Zooplankton türlerinin biyolojik çe?itlili?i, kompozisyonu ve mevsimsel de?i?iklili?i
Sinop bölgesi Bo?azlardan uzakta oldu?undan Akdenizle?menin etkisi az olup burada tan?mlanan tür miktar? da fazla de?ildir. Akdenizle?me süreci Karadeniz kopepod faunas?n?n zenginle?mesinde çok önemli bir yer tutar. Bu süreç Kovalev ve ark. (1998b) taraf?ndan detayl? biçimde çal???lm??t?r. Akdeniz’den orjinlenen 60 kopepod türü saptanm?? ve bunlardan be?inin (Microcalanus pusillus, Aetideus armatus, Euchaeta marina, Metridia lucens, ve Oncaea obscura) 1995-1997 y?llar? aras?nda Karadeniz’in Bo?aziçi bölgesinden toplanan yeni kay?tlar oldu?u belirtilmi?tir (Kovalev ve ark., 1998b).
Ergün (1994), güney Karadeniz’de 1991- 1992 y?llar?nda yapm?? oldu?u çal??mas?nda 5 bask?n kopepod (Calanus ponticus, Acartia clausi, Pseudocalanus elongatus, Centropages kröyeri ve Paracalanus parvus) türünü tan?mlam??t?r. Yine Besiktepe (2001) çal??mas?nda Karadeniz’in güney-bat? k?sm?nda be? bask?n kopepod türünü listelemi? ancak Ergün (1994)’den farkl? olarak Centropages ponticus yerine Oithona similis türünü tespit etmi?tir. Y?ld?z (1997), Karadeniz’in Sinop ilinin ?ç Liman bölgesinde gerçekle?tirmi? oldu?u çal??mas?nda Calanus helgolandicus, Paracalanus parvus, Pseudocalanus elongatus, Acartia clausi ve Oithona nana olmak üzere yine be? kopepod türünü tespit etmi?tir. Ünal (2002), 1999 y?l?nda Sinop bölgesinde gerçekle?tirdi?i çal??mas?nda yedi kopepod türünün (Calanus euxinus, Acartia clausi, Pseudocalanus elongatus, Oithona similis, Centropages ponticus, Acartia tonsa ve Pontella mediterrenea) varl???n? belirlemi?tir. Çal??mam?zda toplam alt? kopepod türü tan?mlanm?? olup, Ünal (2002)’dan farkl? olarak Pontella mediterrenea türüne rastlan?lmam??t?r. Erkan ve ark. (2000), güneydo?u Karadeniz’de zooplanktonun günlük vertikal da??l?m?n? çal??m?? ve Copepoda, Noctiluca, Cladocera, Polychaeta larva, Appendicularia, Bivalvia, Chaetognatha, Ostracoda, Coelenterata, Ctenophora, Tintinnidae ve Gastropod larva gibi ba?l?ca zooplankton gruplar?n?n varl???n? tan?mlam??lard?r.
Mesozooplankton, besin zooplankton ve Noctiluca türünü kapsamaktad?r. Üstün, (2005) taraf?ndan yap?lan çal??mada k?y?sal istasyonda aç?k istasyona oranla mesozooplankton bollu?unun yüksek de?erlere sahip olmas? beklenirken, istasyonlar aras?nda dikkate de?er bir farkl?l?k belirlenmemi?tir (?ekil 2.). Benzer durum Ünal (2002)’?n çal??mas?nda da gözlenmi?tir. Bunun nedeni istasyonlar aras?ndaki mesafenin uzak olmamas? (7.5-9.5 km kadar) ve aç?k istasyondaki su dinami?inden kaynaklanm?? olabilir. Bundan dolay? birçok çal??mada Sinop bölgesi aç?klar? upwelling (nütrient ta??n?m ?ekli) alan? olarak kabul edilmi?tir (Niermann ve ark., 1994; Kideys ve ark., 1999).
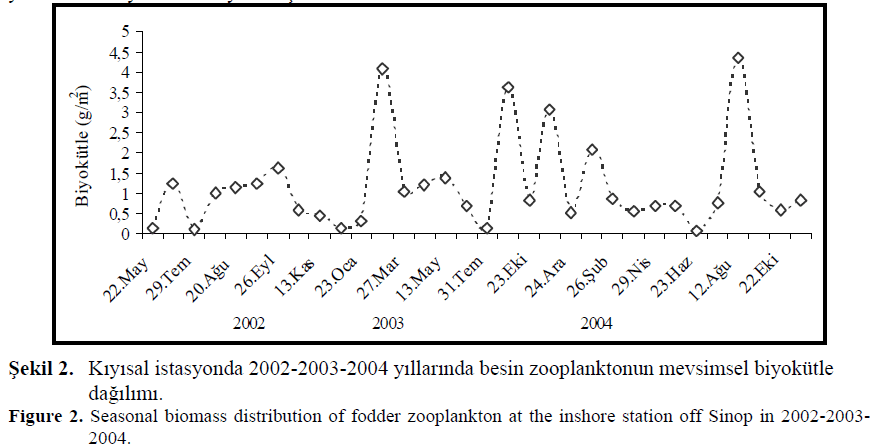
?ekil 2. K?y?sal istasyonda 2002-2003-2004 y?llar?nda besin zooplanktonun mevsimsel biyokütle da??l?m?.
Figure 2. Seasonal biomass distribution of fodder zooplankton at the inshore station off Sinop in 2002-2003-2004.
Ünal (2002), çal??mas?nda mesozooplanktonun y?l boyunca 4 farkl? (k??, ilkbahar,yaz ve sonbahar) yüksek de?er sergiledi?ini saptam??t?r. Sonbaharda elde edilen yüksek de?erin, k?? yüksek de?erinden daha belirgin oldu?unu belirtmi?tir. Üstün (2005) taraf?ndan yap?lan çal??mada mesozooplanktonun bolluk ve biyokütle de?erlerinde, 2002 y?l?nda y?l boyunca düzenli veri elde edilemedi?inden dolay? sadece sonbaharda (Eylül ve Kas?m), 2003 y?l?nda k?? (?ubat), sonbahar (Eylül-Kas?m) ve ilkbaharda (Nisan) ve 2004 y?l?nda ilkbahar, yaz ve sonbahar olmak üzere 3 yüksek de?er tespit edilmi?tir. Ünal (2002)’dan farkl? olarak 2003 y?l?nda k?? yüksek de?erinin sonbahar de?erinden daha belirgin oldu?u bulunmu?tur.
Bat ve ark. (2005) taraf?ndan 2002-2004 y?llar?nda k?y?sal istasyon için ortalama biyokütle de?erleri s?ras?yla 0.8 – 1.5 ve 1.3 g/m2 olarak tespit edilmi?tir. Maksimum biyokütle de?erleri 2002 y?l?nda 26 Eylül (1.6 g/m2), 2003 y?l?nda 25 ?ubat (4.1 g/m2) ve 2004 y?l?nda 12 A?ustos tarihinde (4.3 g/m2) saptanm??t?r (?ekil 2.). Ünal (2002), k?y?sal istasyon için ortalama biyokütle de?erini 2.3 g/m2 olarak bulmu? ve maksimum k?y?sal besin zooplankton biyokütle de?erini de ?ubat ay?nda (7.3 g/m2) belirlemi?tir.
Gubanova ve ark. (2001), 1976-1996 y?llar? boyunca Sevastopol Körfezi’nin ba?l?ca gruplar?n?n ve kopepod türlerinin tür kompozisyon bolluk de?erlerinin (birey/m3) mevsimsel ve y?l içi de?i?imlerini çal??m??lard?r (Tablo 6). Kar??la?t?r?lma yap?lmas? amac? ile Gubanova ve ark. (2001), Ünal (2002) ve Üstün (2005) taraf?ndan yap?lna ara?t?rmalar?n verileri Tablo 6’da gösterilmi?tir. Gubonova ve ark. (2001)’n?n çal??ma alan?n?n derinlikleri, Ünal (2002) ve ara?t?rma alan?m?z?n örnekleme derinliklerinden farkl? oldu?undan dolay? verilerin birim hacim olarak kar??la?t?r?lmas? uygun de?ildir (ortalama derinlik Sevastopol Körfezi’nde 12 m ve Sinop Liman?’nda 50-70 m). Bu yüzden, her iki bölgenin besin ve toplam zooplankton de?erleri için birim alan hesaplamalar? yap?larak tabloya dahil edilmi?tir. Birim alana dayanarak, Ünal (2002)’?n çal??mas?nda elde etti?i toplam bolluk de?erinin (128348 birey/m2) Sevastopol’da 1990’dan bu yana belirlenen de?erlerden (18564 birey/m2) 7 kat daha fazla oldu?u görülmü?tür. Ara?t?rmam?zda 2002 y?l?nda 43158 birey/m2, 2003 y?l?nda 91678 birey/m2 ve 2004 y?l?nda ise 71908 birey/m2 olarak toplam bolluk de?erleri hesaplanm??t?r. Bat ve ark. (2005) taraf?ndan yap?lan çal??mada besin zooplaktonun bolluk de?erleri s?ras?yla, 2002 y?l?nda 29065 birey/m2, 2003 y?l?nda 79978 birey/m2 ve 2004 y?l?nda 38590 birey/m2 olarak bulunmu?tur. Ünal (2002)’?n tespit etti?i besin zooplankton bolluk de?eri (68925 birey/m2) 1990’da Sevastopol Körfezi (14196 birey/m2) ile kar??la?t?r?ld???nda 5 kat artt??? gözlenmi? ve bu durumun kopepod, N. scintillans ve meroplankton gibi gruplar aras?ndaki büyüklük farkl?l???ndan kaynaklanm?? olabilece?ini belirtmi?tir.
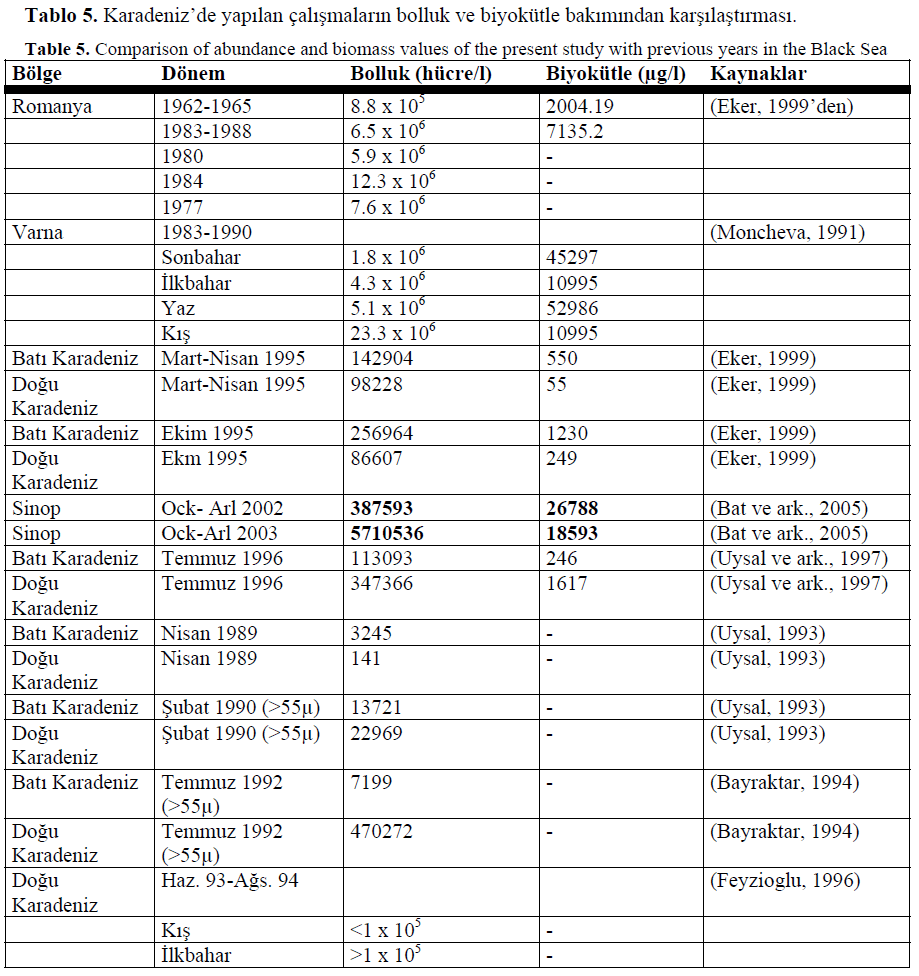
Tablo 5. Karadeniz’de yap?lan çal??malar?n bolluk ve biyokütle bak?m?ndan kar??la?t?rmas?.
Table 5. Comparison of abundance and biomass values of the present study with previous years in the Black Sea
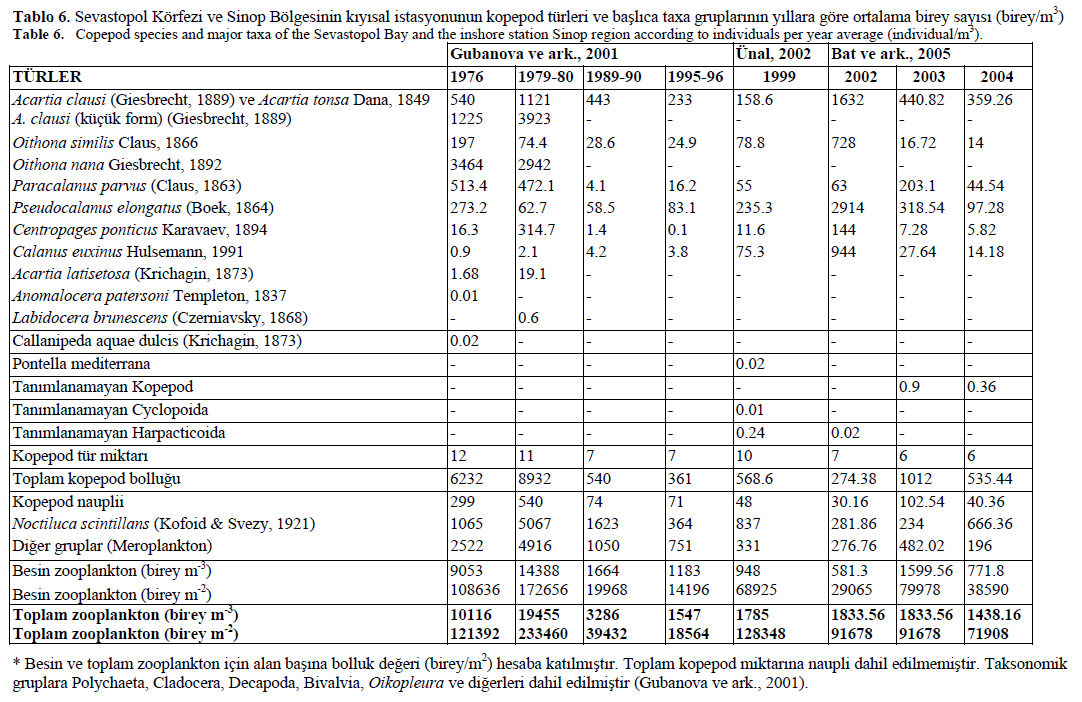
Tablo 6. Sevastopol Körfezi ve Sinop Bölgesinin k?y?sal istasyonunun kopepod türleri ve ba?l?ca taxa gruplar?n?n y?llara göre ortalama birey say?s? (birey/m3)
Table 6. Copepod species and major taxa of the Sevastopol Bay and the inshore station Sinop region according to individuals per year average (individual/m3).
Ara?t?rmalar mevsimsel de?i?iklik ve s?cakl?k aras?nda iyi tan?mlanan ili?kinin bulundu?unu ve bunun yan?nda kalanoid kopepodlar?n mevsimsel dinami?inin habitatlar?na da ba?l? oldu?unu göstermi?tir. Örne?in, Acartia clausi so?uk mevsim boyunca görülürken, Acartia tonsa türünün y?l boyunca sadece ?l?k sularda mevcut oldu?u belirlenmi?tir (Sullivan ve McManus, 1986). A. tonsa türü ABD, Long Island çevresinde sonbahar ve yaz aylar?nda, A. clausi türü k?? ve ilkbahar aylar?nda bol miktarda bulunmu?tur (Conover 1956; Gubanova ve ark., 2001’den). Ünal (2002) çal??mas?nda, Acartia tonsa türünü yaz ortas? ve sonbahar ortas?nda saptam??t?r. ?ubat ve Haziran aylar?nda tespit edememi?tir. En yüksek bolluk de?erini (11092 birey/m2) 2 Kas?m tarihinde gösterdi?ini belirlemi?tir. Acartia tonsa türünün Karadeniz’e 1976’da geldi?i belirlenmi? (Gubanova, 2000); bununla birlikte, Akdeniz’de ilk defa 1985’te kaydedilmi?tir (Belmonte ve ark., 1994).
A. tonsa muhtemelen dünya okyanuslar?n?n baz? bölgelerinden gemilerin balast sular? ile ta??nm??t?r. Karadeniz’in Sinop bölgesinde ve Sevastopol Körfezi’nde Acartia tonsa türünün mevsimsel da??l?m?n?n kontrolünde s?cakl?k önemli bir faktör olmu?tur. Sevastopol Körfezi’nde ?ubattan May?sa kadar tespit edilememi? (Gubanova ve ark., 2001) ancak su s?cakl??? 16ºC’ye yükseldi?inde May?s sonuna do?ru erginleri ilk defa görülmü?tür. A. tonsa türünün bollu?u deniz suyu s?cakl??? 22ºC’ye yükseldi?i zaman Haziran sonundan A?ustos ay?na kadar A. clausi türünü a?m??t?r. En yüksek de?eri A?ustosta (1232 birey/m3) kaydedilmi?tir. Bununla birlikte, deniz suyu s?cakl???n?n azalmas?yla, A. tonsa türünün bollu?u düzenli olarak Eylül’de metre küpte 7 kopepoda kadar azalm??t?r (Gubanova ve ark., 2001). Çal??mam?zda A. tonsa türü 2002 y?l?nda her iki istasyonda da sadece 26 Eylül tarihinde (900 birey/m2) bulunmu?, 2003 ve 2004 y?llar?nda ise bu tür tespit edilememi?tir. Bu tarihte s?cakl?k de?eri 25°C olarak ölçülmü?tür.
A. clausi türünün öritermal tür olup geni? s?cakl?k tolerans?na sahip oldu?u bilinmektedir (Berdnikov ve ark., 1999). Üstün (2005) taraf?ndan yap?lan çal??mada 26 Eylül 2002 tarihinde (36000 birey/m2), 2003 y?l?nda ise Kas?m ay?nda (93500 birey/m2) ve 2004 y?l?nda A?ustos ay?nda (61600 birey/m2) oldukça bol miktarda bulunmu?tur. Ünal (2002), çal??mas?nda Nisan ay?nda en yüksek k?y?sal bolluk de?erini (46095 birey/m2) tespit etmi?tir. Gubanova ve ark. (2001), Sevastopol Körfezi’nde A. clausi türünün Haziran ve Eylül aylar?nda en yüksek de?ere sahip olduklar?n? ve ergin bireylerinin tüm y?l boyunca bulundu?unu kaydetmi?lerdir (177 ve 277 birey/m3).
Gubanova ve ark. (2001), 3 farkl? faktörün etkile?imine bakarak son yirmi y?lda (Beroe türünün geli?inden önce) zooplanktonun dü?ü?ü için sebeplerin neler olacabilece?ini belirtmi?lerdir. Birincisini 1978 boyunca yap?lan dalgak?ranlar taraf?ndan desteklenen evsel at?klar?n bo?alt?m? olu?turmu?tur. ?kinci faktör ise Karadeniz’e balast sular? ile ta??nan Mnemiopsis leidyi olmu?tur. Sonuncusunu ise, Kuzey Atlantik Sirkülasyonun de?i?ikli?inin, 1980’lerin sonu 1990’lar?n ba??nda kuzey yar?m kürede plankton üzerinde (ço?unluklu birey miktar?ndaki azal???) etkiye sahip oldu?u hipotezinin göz önünde tutulmas? meydana getirmi?tir (Niermann ve ark., 1999). Acartia clausi, A. tonsa, A. clausi (küçük from) ve dinoflagellat Noctiluca scintillans gibi ötrofik türler Sevastopol Körfezi’nin artan ötrofikasyonu taraf?ndan tercih edilen türler olmu?lard?r. Di?er taraftan, 1980’de Mart ay?ndan Kas?ma kadar 18.5°C ortalama s?cakl?k, 1976’da 16°C ortalama s?cakl?k ile kar??la?t?r?l?nca Acartia tonsa gibi ?l?k su kopepodlar?n?n miktar?n?n art?? gösterdi?i öne sürülmü?tür (Gubanova ve ark., 2001).
?htiyoplankton
Besin zincirinin ilk halkas?n? olu?turan birincil üreticiler ve tüketiciler olarak adland?rd???m?z plankton, su içerisinde aktif hareket yetene?inden yoksun olup su hareketleriyle ta??nan canl?lar?n (organizmalar?n) s?n?fland?r?lmas?nda da kullan?lmaktad?r. Bu gruba denizde ya?ayan hemen her taksonomik grup girmektedir. Bal?klar?n ise yumurta ve larvalar?n? k?smen de genç bireylerini içeren grubu, ihtiyoplankton yani bal?k planktonu olarak adland?r?l?r. ?htiyoplanktonoloji, ihtiyoloji biliminden, bal?k biyolojisine ve hatta akuakültür ara?t?rmalar?na kadar bütün ilgili bilim dallar? aç?s?ndan son derece önemlidir. Ayr?ca bal?klar?n insan besini olarak kullan?lmas? nedeniyle stoklar?n yönetimi aç?s?ndan da özel bir öneme sahiptir. Bu tip çal??malar; bal?klar?n yumurtlama dönemlerini, alanlar?n? ve bunlarda görülen zamansal de?i?imleri saptamak, yumurtlayan ergin sto?un büyüklü?ünü tahmin etmek, herhangi bir türün, bir yumurtlama dönemi sonucunda olu?acak y?l s?n?f?n?n ya?am oran?n? ya da di?er bir deyi?le ölüm oran?n? tahmin etmek ve buna etki eden faktörleri irdelemek, ihtiyoplanktonun fizyolojik, morfolojik, genetik, taksonomik ve çevre ile olan ili?kilerini incelemek aç?s?ndan gereklidir (Yüksek ve Gücü, 1994).
Bu amaçlar do?rultusunda yap?lan çal??malar?n ba??nda bal?klar?n ne zaman ve nerede yumurta b?rakt?klar?, söz konusu alanlarda yumurtlama, embriyo ve larval geli?me-beslenme s?ras?ndaki çevresel ko?ullar?n belirlenmesi, di?er çal??malar için bir ön etüt niteli?inde olup, son derece önemlidir (Yüksek ve Gücü, 1994). Hangi tür veya türlerin ne zaman, nerede ve hangi ko?ullarda yumurta b?rakt??? bilindi?i taktirde üreme davran??lar?, yumurtlama, embriyonik ve larval geli?me evrelerindeki kay?plar, bunlarda meydana gelen zamansal de?i?im ile dalgalanmalar ve ergin stoka kat?l?m oranlar? tahmin edilebilir (Yüksek, 1993).
Karadeniz’in baz? türler hariç kendine özgü bir faunas? yoktur. Genelde Akdeniz-Atlantik formlar? taraf?ndan i?gal edilmi?tir. Sucul ortamda bulunan canl?lar do?al bir dengede varl?klar?n? sürdürürler. Son y?llarda bu ekolojik denge de?i?ime u?ram??t?r. Bu de?i?imden yararlanan birçok tür bu ortamda hakimiyet sa?lamaya ba?lam??t?r. Son olarak Jelimsi makrozooplankton ekosisteme girmi? ve tamamen sistemi etkilemi?tir (Vladimirov ve ark., 1999).
Karadeniz Ekosisteminde Meydana De?i?imler ve Etkileri
Karadeniz bal?kç?l???n?n Türkiye’nin canl? deniz kaynaklar? içerisinde özel bir yeri vard?r. Karadeniz’de avlanan bal?klar toplam üretimin %82’sini olu?turmaktad?r. Türkiye, hamside a??r? avc?l???n etkilerini bal?kç?l?k ekipmanlar?n?n geli?imi ile 1987-1988 bal?k avc?l??? sezonunda ya?am??t?r. Bunun yan?nda mesozooplankton biyokütlesi M. leidyi taraf?ndan kullan?lm?? ve sonucunda ktenofor kitlesinin a??r? art??? ile kar??la??lm??t?r. Mesozooplankton miktar?n?n azalmas? küçük pelajik bal?k miktar?nda de?i?imler meydana getirmi?tir. Bal?klar?n beslenmesinde zooplankton yetersizli?i ilk olarak bal?k boylar?nda küçülmeye ve daha sonraki y?llarda stoklar?n çökü?üne neden olmu?tur (Niermann ve ark., 1999; Gucu, 2002). Karadeniz’in Türkiye k?y?lar?ndaki hamsi av miktar?n?n 1988 y?l?nda 295000 ton oldu?u 1989 y?l?nda 97.000 tona geriledi?i belirlenmi?tir. 1990’da daha da dü?en av, 66000 tona gerilemi?tir. 1990 y?l?ndan sonraki y?llarda h?zl? bir art???n söz konusu oldu?u 1992 y?l?nda 155000 ton, 1995 y?l?nda 373.000 ton seviyesine ula?t??? görülmektedir (?ekil 3., Anonim, 2004).
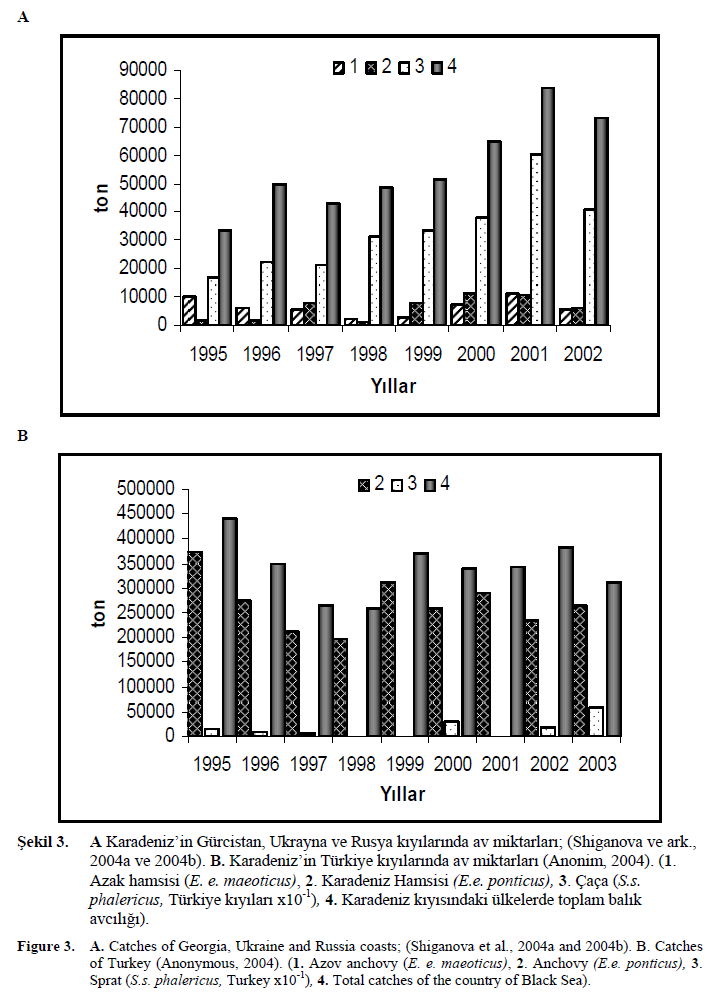
?ekil 3. A Karadeniz’in Gürcistan, Ukrayna ve Rusya k?y?lar?nda av miktarlar?; (Shiganova ve ark., 2004a ve 2004b). B. Karadeniz’in Türkiye k?y?lar?nda av miktarlar? (Anonim, 2004). (1. Azak hamsisi (E. e. maeoticus), 2. Karadeniz Hamsisi (E.e. ponticus), 3. Çaça (S.s. phalericus, Türkiye k?y?lar? x10-1), 4. Karadeniz k?y?s?ndaki ülkelerde toplam bal?k avc?l???).
Figure 3. A. Catches of Georgia, Ukraine and Russia coasts; (Shiganova et al., 2004a and 2004b). B. Catches of Turkey (Anonymous, 2004). (1. Azov anchovy (E. e. maeoticus), 2. Anchovy (E.e. ponticus), 3. Sprat (S.s. phalericus, Turkey x10-1), 4. Total catches of the country of Black Sea).
Karadeniz’e k?y?s? olan di?er ülkelerden Gürcistan, Rusya ve Ukrayna’da toplam bal?k avc?l??? miktar?nda 1995 y?l?nda dü?ü? kaydedilmi?tir. 2000 y?l?ndan itibaren ise daha önce iyice azalan avc?l?k miktar? yükselmeye ba?lam??t?r (?ekil 3). Beroe Karadeniz’e girmeden önceki y?llarda Rusya k?y?lar?nda Karadeniz hamsisi tamam?yla yok olmu?ken 2000 y?l?ndaki avc?l?k sezonunda tekrar avlanmaya ba?lam??t?r (Shiganova, 2004; Shiganova ve ark., 2004a). Bu ülkelerin 1995 y?l?nda av miktarlar? dü?mü? iken Karadeniz’in Türkiye k?y?lar?nda art?? görülmü?tür. Bu farkl?l???n nedeni M. leidyi geli?inden sonra hamsinin yumurtlama alan?n?n Güneydo?u Karadeniz’e kaymas? olarak gösterilmektedir (Shiganova ve ark., 2004a ve 2004b). Fakat Türkiye k?y?lar?nda daha sonraki y?llarda azalmaya ba?layan av miktar?, hamsi, çaça gibi pelajik türler ve toplam bal?k avc?l??? 1998 y?l?ndan sonra dalgal? bir art?? göstermi?tir (?ekil 3).
Karadeniz’de k?y?s? olan ülkelerde genel olarak av miktar?nda 2000 y?llar?nda art?? olurken benzer olarak M. leidyi miktar?ndaki art?? 2000’li y?llarda dü?ü? izlemi?tir; avc?l?k ve M. leidyi türü aras?ndaki bu paralellik dikkat çekicidir (Anonim, 2004).
Karadeniz ekosistemindeki planktonun genel ve özel olarak uzun dönem de?i?iklikleri su rezervi üzerine insan kaynakl? etkiyle ayn? zamana rastlar. K?y?sal kuzey-bat? Karadeniz için bütün temel planktonik bile?enlerin tür kompozisyonu ve miktar?ndaki de?i?iklikler ötrofikasyon, kirlilik ve ayr?ca istilac? M. leidyi türü ile ili?kili oldu?u görülmektedir (Vinogradov ve Flint, 1985; Kovalev ve ark., 1993; Shiganova ve ark., 2001 ve Shiganova, 2004). Karadeniz planktonundaki uzun dönem de?i?iklikler hidro-meteorolojik durumlardaki de?i?melerden kaynaklanabilir (Zhorov ve Boguslavsky, 1985; Vinogradov, 1987; Niermann ve ark., 1998).
Kovalev ve ark. (1998a) Karadeniz’in uzun dönem zooplankton yap?s?n? incelemi?ler ve 1950’nin sonundan 1960’lar?n ortas?na kadar kuzey-bat? k?y? bölgesinin biyokütlesi kuzeydo?u bölgesininkinden yakla??k 3 kat daha yüksek ç?km??t?r. 1970’lerde her iki alan?n biyokütlesi ayn? seviyeye gelmi?tir. Kuzey-bat? k?y? bölgesinde 1980’nin sonundan 1990’n?n ba??na kadar az ya da çok stabil iken, 1990’a kadar kuzey-do?udaki zooplankton biyokütlesi artm??t?r. Sonra her iki alan için biyokütle de?eri önceki y?llardaki ortalaman?n da alt?na dü?mü?tür. 1988’den beri Kuzey-bat? k?y? bölgesinin biyokütlesi Kuzeydo?ununkinden daha dü?ük bulunmu?, tersi durum 1960’da görülmü?tür. Kuzey-bat? Karadeniz’de yem (mesezooplankton, Noctiluca ve jelimsi organizmlar hariç) zooplanktonun son 30 y?l boyunca azalmas?na kar??l?k Kuzey-do?u Karadeniz zooplanktonunun biyokütlesi 1990’a kadar artm??t?r. Her iki bölgede biyokütle 1993’den sonra artm??, 1995’de maksimum seviyeye ula?m??t?r. Güney-do?u Karadeniz aç?klar?ndaki biyokütle güney-bat? aç?klar?ndan daha h?zl? artm?? ve 1994-1996 periyodu boyunca güney-do?u alan?nda yakla??k 2 kez biyokütle art??? gözlenmi?tir.
Karadeniz’de jelimsi organizmalar?n besin a??ndaki yeri incelendi?inde gruplar aras?ndaki ili?kiler daha iyi aç?klanabilmektedir. Fitoplankton ile ba?layan beslenme a?? yüksek yap?daki büyük pelajik bal?klar ve yunuslar ile son bulmaktad?r. Jelimsi organizmalar?n birincil olarak etkiledikleri zooplankton, bal?k yumurta ve larvalar?d?r (Shiganova ve ark., 1998 ve 2000a). Karadeniz’de B. ovata türü M. leidyi ve P. pileus türleri ile beslendi?inden bu organizmalar?n da??l?m?n? ve miktar?n? do?rudan etkilemektedir (Shiganova ve ark., 2000b). Beslenme a??nda bulunan küçük pelajik bal?klar (hamsi, çaça gibi) zooplankton ile beslendiklerinden jelimsi organizmalar ile besinlerini payla?mak zorunda kalmaktad?r. Hem zooplankton miktar?n?n azalmas? hemde jelimsi organizmalar?n bal?k yumurta ve larvalar? ile beslenmeleri küçük pelajik bal?klar bu a? içerisinde ikincil olarak etkilenen grubu olu?turmaktad?r. Di?er basamaklarda bulunan canl? gruplar? ise direk veya dolayl? olarak de?i?imlerden etkilenmektedir (Shiganova ve ark., 2004a ve 2004b, ?ekil 4).
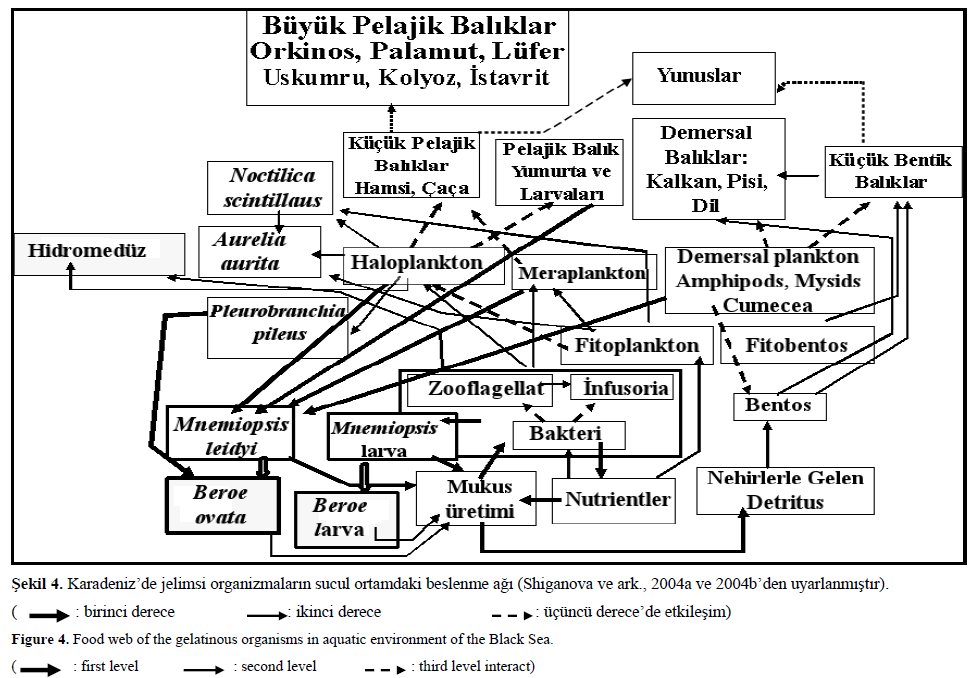
?ekil 4. Karadeniz’de jelimsi organizmalar?n sucul ortamdaki beslenme a?? (Shiganova ve ark., 2004a ve 2004b’den uyarlanm??t?r).
(→: birinci derece →: ikinci derece →: üçüncü derece’de etkile?im)
Figure 4. Food web of the gelatinous organisms in aquatic environment of the Black Sea.
(→: first level →: second level →: third level interact)
Sinop k?y?lar?nda pelajik ekosistemin yap?s?, burada belirgin olarak görülen upwelling ve downwelling olaylar? ile özellik arz etmektedir. Karadeniz’de en önemli ve belirgin upwelling sahas? Tuna, Dinyeper ve Dinyester nehirlerinin bo?ald??? kuzeybat? k?ta sahanl??? ve nispeten K?r?m Yar?madas?’n?n güneyindeki bölgedir. Kuzeybat? Karadeniz d???nda periyodik upwelling olay?n?n görüldü?ü en önemli alan Güneybat? Karadeniz’in bat? k?y?lar?d?r. Bir tür yüzey ak?nt?s? olan upwelling belirli bir hacim içerisindeki suyun, dikey hareketlerle yüzeye ta??nmas? ve yüzeyde yatay yönde nakledilmesi ?eklinde aç?klanabilir. Okyanuslar?n ekvator ku?a?? boyunca yüzey sular?n?n bat?-do?u yönünde esen rüzgarlar?n tesiri ile kuzey yar?m kürede kuzeye ve güney yar?m kürede ise güneye do?ru ta??nmas? sonucu meydana gelen upwelling olay?na “Ekvatoral upweling”, okyanuslar?n ve denizlerin k?ta kenarlar?nda meydana gelen upwellinge de “K?y?sal Upwelling” ad? verilir. Upwelling olay?n?n meydana gelmesine sebep olan faktörler nehir ak?nt?lar?, mevcut ak?nt?lar?n deniz dibi engeline rastlanmas? sonucu yüzeye ta??n?m?, gel-git ak?nt?lar? ve rüzgar tesiri ile meydana gelen yüzey ak?nt?lar?d?r. Tesir bak?m?ndan en önemlisi rüzgar tesiri ile meydana gelen upwelling sirkülasyonudur (Brink, 1978).
Etkin bir upwelling sahas? olan kuzeybat? k?ta sahanl???na ait, özellikle 1980 öncesi y?llarda tespit edilen özelikler, günümüz için geçerli olmayabilir. Çünkü 1990’l? y?llar?n sonuna kadar Karadeniz’e nehirler yoluyla ta??nan nutrient ve kirleticilerin miktar?nda a??r? derecede art?? meydana gelmi?tir (Anonim, 1997). Nutrientteki art???n ana kayna??n? Tuna nehri te?kil etmektedir. Tuna nehri vas?tas?yla ta??nan nutrient ve detritus miktar?ndaki art?? upwelling sirkülasyonunun da tesiriyle havzan?n s?? kuzeybat? sahanl???n?n ekolojik yap?s?n?n bozulmas?na sebep olmaktad?r. Kuzeybat? k?ta sahanl??? sular?nda, özelikle 1970’li y?llardan sonra ortaya ç?kan ötrofik yap? ve bunun sonucu olarak da zemin ve zemin üzerindeki tabakalarda hipoksia ve anoksia olu?umu tespit edilmeye ba?lanm??t?r. Hipoksia ve anoksia olaylar?n?n sonucu ortamda kütlesel ölümler olmaktad?r. Özellikle upwelling periyodunda, zeminden yüzey tabakas?na ta??nan anoksik ve H2S ihtiva eden sular bütün su kütlesinin etkileyerek bölgede kütlesel ölümlere sebep olmaktad?r. Bunun sonucu olarak bölgede ya?ayan bal?k türlerinin toplam stoklar?nda belirgin bir azalman?n meydana geldi?i çe?itli ara?t?r?c?lar taraf?ndan belirtilmektedir. Ayr?ca pek çok tür bölgeyi terk etmek zorunda kalm??t?r. Bu türlerin besin zincirindeki yerinin M. leidyi gibi f?rsatç? türler alm??t?r. Kuzeybat? k?ta sahanl???nda görülen, nutrient konsantrasyonlar?nda ve biyolojik yap?daki de?i?im sadece bu bölgeye özgü olmay?p Karadeniz havzas?n?n her taraf?nda benzer bir de?i?imin oldu?u çe?itli ara?t?r?c?lar taraf?ndan belirtilmektedir. Yo?un ötrofikasyon sonucu özellikle Bat? Karadeniz’de ???k geçirgenli?i oldukça azalm?? ve seki disk derinli?i 1920’lerde 20 m iken 1990’l? y?llarda 5-6 m’ye dü?mü?tür (Sur ve ark., 1993).
Karadeniz’de 1970’lerde alt besin seviyesi durumunda olan nutrientlerin, daha sonralar? fazla miktarda ço?almas? ile beraber ötrofikasyonun etkisi artmaya ba?lam??t?r. Plankton komposizyonunun yap?s?, yo?unlu?u, tür çe?itlili?i ve yatay paralellikte a??r? alg art??lar? de?i?irken, pelajik bal?klar?n besinini olu?turan mesozooplankton gruplar? küçülmü? ve de?erleri azalm??t?r. Bunlarla beraber f?rsatç? ve jelimsi organizmalarda da de?i?imler olmu?tur. Ötrofikasyon ile sars?lan ekosistem daha sonra a??r? avc?l?k ile kar??la?m??t?r. 1970’lerin ba??nda sistemde, geni? boyuttaki pelajik bolluk için küçük pelajik bal?klar ana predatör olmu?lard?r. Mesozooplankton üzerinde güçlü bir grazing etkisi ba?lam?? ve bu nedenle fitoplankton miktar? artm??t?r. A??r? avc?l???n gerçek etkisi küçük pelajik stoklar?n?n 1980’li y?llarda a??r? tüketildi?i anla??ld???nda ortaya ç?km??t?r. Sistemde meydana gelen bo?luktan jelimsi organizmalar ile f?rsatç? türler yararlanm?? ve 1980’lerin sonunda ekosistemin tümünü kontrol etmeye ba?lam??lard?r (Vinogradov ve Shushkina, 1992; Oguz, 2005a).
Karadeniz’de upwelling olay?n?n meydana geldi?i Kuzeybat? k?ta sahanl??? bu duruma iyi bir örnek te?kil eder. Kuzeybat? k?ta sahanl???nda yap?lan çal??malarda; baz? bal?klar?n (çaça, hamsi, kalkan vb.) mevsimsel piknoklin tabakas?n?n alt?ndaki so?uk sularda yumurtlad?klar? tespit edilmi?tir. Upwelling sirkülasyonu sonucu dip sular?n?n yüzeye ta??nmas? esnas?nda bu sulara b?rak?lan yumurtalarda güne? ?????n?n etkili oldu?u yüzey tabakas?na ta??n?r ve yüzeye ta??nan yumurtalardan yavru ç?k?? oran? dip sulara göre büyük oranda artar. Dolay?s?yla hamsi, çaça ve kalkan gibi so?uk su ortam?nda yumurta b?rakan türler, sürekli upwelling olay?n?n meydana geldi?i aylarda (Haziran-A?ustos aras? yaz aylar?nda) normalden daha yüksek oranda yumurtlama yapar ve yumurtadan yavru ç?k?? oran? normalden daha yüksek orandad?r. Ayn? sahada fitoplankton ve zooplanktonun yüksek verimlilikteki geli?imi plankton ile beslenen hamsi ve çaça gibi bal?klar için verimli bir beslenme alan? sa?lar ve bunun sonucu olarak da bölgede yap?lan avc?l?kta yüksek av verimi sa?lanabilmektedir (Erüz ve ark., 1993).
Hamsi
Göçmen bir bal?k olan hamsi yakla??k 3-4 y?l ya?ar. E?eysel olgunlu?a ula?ma birinci y?l?n sonunda olur. Yani yumurtadan ç?kan hamsi larvalar? 11-12 ay sonra cinsi olgunlu?a eri?ir ve ya?am boyunca iki defa döl verir. Her yumurtlama periyodunda 13.000-40.000 civar?nda yumurta b?rak?r. Hamsi bal???nda üreme genel olarak su s?cakl???n?n 17.5-27°C, tuzlulu?un ‰ 12-18, pH’n?n 8.3-8.4 ve derinli?in 5- 10 m oldu?u k?y? bölgelerinde meydana gelir (Slastenenko, 1955-1956).
Hamsinin yumurtlama ve beslenme alanlar?, Karadeniz’in kuzeybat? bölgeleridir. Bu bölgelerde yaz?n beslenen ve yumurtlayan hamsiler, k??lamak için daha ?l?man olan Karadeniz’in Türkiye sahillerine göç ederler. Ancak son y?llarda hamsi stoklar?n?n özellikle geleneksel üreme alanlar?nda baz? de?i?imler oldu?u ileri sürülmektedir. Niermann ve ark. (1994) taraf?ndan 1991 ve 1992 y?llar?nda yap?lan çal??mada, hamsinin kuzeybat? Karadeniz’deki geleneksel yumurtlama alanlar?n? terk ederek güneydo?u Karadeniz k?y?lar?n? yeni yumurtlama alanlar? olarak seçti?i tespit edilmi?tir.
Genel olarak hamsi yumurta ve larvalar?na Sinop k?y?lar?nda May?s-Eylül aylar?nda rastlanm?? olup (Sat?lm??, 2001 ve 2005), vertikal çekimlerde en yo?un yumurta ve larva Temmuz’da bulunmu?, horizontal çekimlerde ise yumurta yo?unlu?una Temmuz’da, larva yo?unlu?una ise Haziran’da rastlanm??t?r (Sat?lm??, 2005). 24 saatlik kuluçka süresi dikkate al?nd???nda yumurta ile larva aras?nda bir s?ralama olmamas? normal olarak tan?mlanabilir. Hamsilerin yumurtlama zamanlar?na ili?kin çe?itli ara?t?rmac?lar?n farkl? sularda bulduklar? sonuçlar a?a??daki çizelgede verilmi?tir (Tablo 7.).
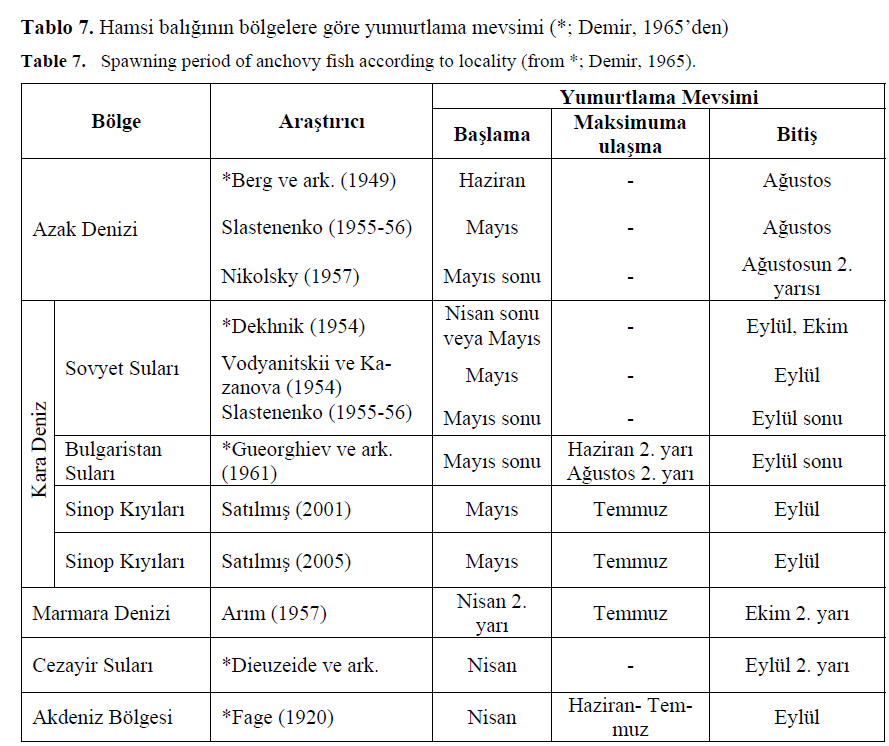
Tablo 7. Hamsi bal???n?n bölgelere göre yumurtlama mevsimi (*; Demir, 1965’den)
Table 7. Spawning period of anchovy fish according to locality (from *; Demir, 1965).
Tablo 7.’e göre hamsilerde yumurtlama Azak Denizi’nde May?s, May?s sonu ve Haziran’da ba?lay?p A?ustos’ta son bulurken, Karadeniz’de May?s’ta ba?lay?p Eylül’de, Marmara ve Akdeniz’de Nisan’da ba?lay?p Eylül’de sona ermektedir. Mevcut ara?t?rmada ve bu bölgede daha önce yap?lan ara?t?rmada bulunan May?s-Eylül aras? yumurtlama mevsimi di?er ara?t?r?c?lar taraf?ndan da ayn? bulunmu?tur. Demir (1965), hamsilerde yumurtlaman?n Nisan ay?ndan önce ba?lamad???n? ve Kas?m ay?ndan sonra da devam etmedi?ini (ço?unlu?u Ekim’den sonra) ve yumurtlaman?n yaz aylar?nda maksimuma ula?t???n? belirtmi?tir.
Vertikal çekimlerde elde edilen yumurtalar 7.5-260 adet/m2, larvalar ise 5-115 adet/m2 bollukta, horizontal çekimlerde ise yumurtalar kaydedilmi?tir. A?ustos 1993 sürekli olarak dü?ük say?da yumurta kaydedilmi? ve A?ustos, yumurtlama mevsiminin sonuna tekabül eden ay olarak aç?klanm??t?r (Slastenenko, 1955/1956). Sat?lm?? (2005) taraf?nda yap?lan çal??mada ise di?er ara?t?rmalara göre yumur ve larvada en yüksek de?ere ula??lm??t?r. Son y?llarda hamsi yumurta ve larva say?lar?ndaki art?? e?ilimi Karadeniz ekosistemininin düzelmesinin bir i?areti olarak yorumlanabilir.
Gordina ve ark. (2005), hamsi yumurtalar?n? 2000 y?l? Haziran-A?ustos döneminde vertikal örneklemelerde Ukrayna sular?nda 10.2 adet/m2, Sinop Bölgesinde 31.7 adet/m2, horizontal örneklemelerde Ukrayna sular?nda 62.5 adet/100m3, Sinop bölgesinde 104.6 adet/100m3, 2001 y?l? Haziran-A?ustos döneminde vertikalde 31,6 adet/m2, horizontalde 116.6 adet/100m3 olarak tespit etmi?lerdir. Larvalar?n? ise 2000 y?l? Haziran- A?ustos döneminde vertikal örneklemelerde Ukrayna sular?nda 0.7 adet/m2, Sinop bölgesinde 2.5 adet/m2, horizontal örneklemelerde Ukrayna sular?nda 0.9 adet/100m3, Sinop bölgesinde 0.2 adet/100m3, 2001 y?l? Haziran-A?ustos döneminde vertikalde 1.2 adet/m2, horizontalde 0.9 adet/100m3 olarak bulmu?lard?r. 2000 y?l? dikkate al?nd???nda hamsi yumurtalar? Sinop bölgesinde Ukrayna sular?ndan vertikal çekimlerde 3 kat, horziontal çekimlerde 1.7 kat daha fazla bulunurken, larvalar? Sinop bölgesinde vertikal çekimlerde Ukrayna sular?ndan 3,5 kat fazla bulunurken, horiziontal çekimlerde 4.5 kat az bulunmu?tur. Sat?lm?? (2005) taraf?ndan yap?lan çal??mada Haziran- Temmuz 2003 döneminde Sinop bölgesinde yumurta miktar? vertikalde 182 (±58.26) adet/m2, horizontalde 42.63 (±16.70) adet/100m3, larva ise vertikalde 49 (±16.50) adet/m2, horizontalde 11.75 (±5.68) adet/100m3 bollukta hesaplanm??t?r. Bu sonuçlar daha önce Sinop ve Ukrayna’da yap?lan ara?t?rmalarla k?yasland???nda yumurtalar?n vertikalde di?er ara?t?rmalar?n sonuçlar?ndan fazla bulunurken, horizontalde daha az; larvalarda ise hem vertikalde hem de horizontalde di?er ara?t?rma sonuçlar?ndan daha fazla bulunmu?tur.
Hamsi yumurtalar?n?n en yo?un bulundu?u Haziran ve Temmuz aylar?nda s?ras?yla, s?cakl?k 16.5-24oC, 23oC, tuzluluk ‰ 17.4- 17.9, ‰ 17.9, pH 8.47-8.63, oksijen 7.7-12.1 mg/l, 7.4-12.3 mg/l olarak tespit edilmi?tir (Sat?lm??, 2005). Niermann ve ark. (1994), yüzey (5 m) suyu s?cakl?klar?n? Haziran 1991’de 16 oC, Do?u Karadeniz’de 17.8-19oC, Temmuz 1992’de 20.2-22.5oC, yine Do?u Karadeniz’de 23-24oC aras?nda oldu?unu bulmu?lard?r. Hamsi yumurtalar?n?n olgunla?maya ba?lad??? minimum s?cakl???n 13 oC, yumurtlaman?n ba?lad??? s?cakl?k aral???n?n ise 13/14-26 oC aras?nda oldu?unu, maksimum yumurtlaman?n 20oC’den daha yüksek s?cakl?klarda oldu?u; yumurtlaman?n oldu?u suyun tuzlulu?unun ‰ 12-18 ve pH’s?n?n 8.3- 8.4 aras?nda de?i?ti?i bildirilmektedir (Demir, 1959).
Pavloskaya (1955), bir di?i Karadeniz hamsisi taraf?ndan üretilen yumurtalar?n %25- 35 oran?nda ya?ama oran?na sahip oldu?unu, larva ve juvenil hamsilerin ya?ama oranlar?n?n ise özellikle g?da olarak bulunacak planktonun miktar ve kalitesine ba?l? oldu?unu rapor etmi?tir. Karnilova (1955) 1950-53 y?l?nda yumurtlama sezonunda Azak Denizi’nin 0-1 m derinli?inde ald??? 1 m3’lük su örneklerinde ya?ama oran?n? yumurtalar için %0.1-9.6 aras?nda bulmu?tur. Gordina ve ark. (1990) genellikle May?sta b?rak?lan (erken parti) yumurtalardan ç?kan larvalarda yüksek ölüm oranlar? görüldü?ünü belirtmi?lerdir. Bu durumun larvalar?n dikey göç s?ras?nda so?uk su ile kar??la?malar?ndan kaynakland???n?, en yüksek ya?ama oran?n?n Haziran sonu- Temmuz ba??nda b?rak?lan yumurtalarda görüldü?ünü aç?klam??lard?r.
Karadeniz hamsi stoklar? üzerinde çal??ma yapan de?i?ik ara?t?r?c?lar taraf?ndan farkl? y?llarda bulunan ortalama boy-a??rl?k, cinsiyet oranlar? Tablo 9’da verilmi?tir.
Erkoyuncu ve Özdamar (1989), 1986 y?l?nda yapt?klar? ara?t?rmada ortalama a??rl??? 10.5 g olarak bulmu?lard?r. Bu y?ldan itibaren yap?lan çal??malarda ayn? tür a?larla avlanan hamsilerin ortalama a??rl?klar?nda sürekli bir dü?me e?ilimi görülmü? ve 1991 y?l?nda 3.9 g olarak hesaplanm??t?r. 1989-1990 av sezonundan itibaren yürürlü?e konan 9.0 cm’den küçük boydaki hamsilerin avlanmamas? sebebiyle avlanan bireyler içindeki büyük ya? gruplar?na ait bireylerin miktarlar?nda art??lar olmu?, dolay?s?yla ortalama a??rl?klar da 1992 y?l?ndan itibaren tekrar yükselmeye ba?lam?? ve 1997- 98 av sezonunda 8.67 g’a ula?m?? (Gözler ve Çilo?lu, 1998), Sat?lm?? (2005) taraf?ndan yap?lan çal??mada ise bu çal??madan biraz daha küçük 7.8 g olarak bulunmu?tur (Tablo 9).0.4- 159 adet/100m3, larvalar ise 2.6-52.1 adet/100m3 aras?nda de?i?en bolluk de?erlerinde bulunmu?tur (Sat?lm??, 2005).
Niermann ve ark. (1994) 1991 y?l?nda hamsi yumurtalar?na k?y?sal sularda 10-30 adet/m2, aç?k sularda ise 0-2 adet/m2 bulmu?lar Haziran 1991 y?l?nda yapt?klar? bu çal??mada Türkiye k?y?lar?nda 66 istasyonda 154 yumurta, Ukrayna k?y?lar?nda ise yine 66 istasyonda 286 yumurta bulmu?lard?r. Larvaya ise sadece iki istasyonda 2 adet/m2 larvaya rastlam??lard?r. Temmuz 1992’de ise Sinop aç?klar?nda yumurtay? 0-40 adet/m2, larvay? ise maksimum 26 adet/m2 olarak tespit etmi?lerdir. Güney Karadeniz’de yap?lan farkl? hamsi yumurta ve larva sörveylerine ili?kin sonuçlar a?a??daki Tablo 8’de verilmi?tir.
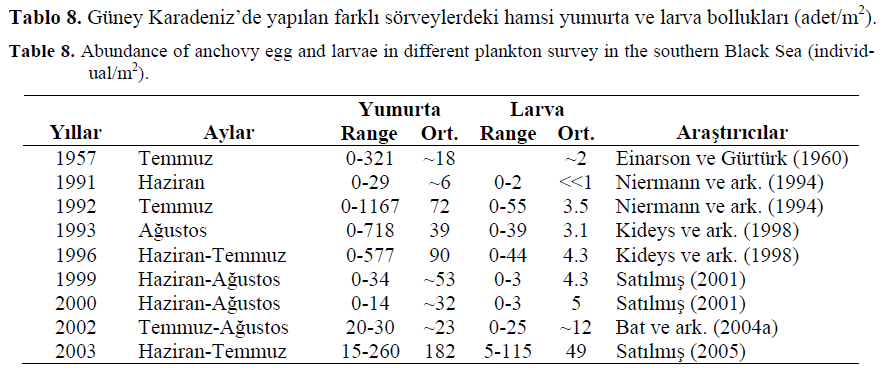
Tablo 8. Güney Karadeniz’de yap?lan farkl? sörveylerdeki hamsi yumurta ve larva bolluklar? (adet/m2).
Table 8. Abundance of anchovy egg and larvae in different plankton survey in the southern Black Sea (individual/m2).
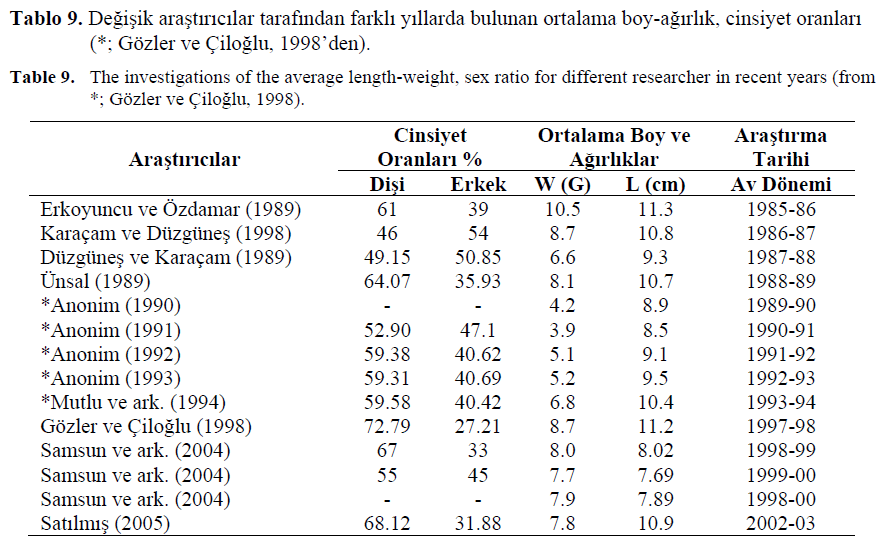
Tablo 9. De?i?ik ara?t?r?c?lar taraf?ndan farkl? y?llarda bulunan ortalama boy-a??rl?k, cinsiyet oranlar?
(*; Gözler ve Çilo?lu, 1998’den).
Table 9. The investigations of the average length-weight, sex ratio for different researcher in recent years (from
*; Gözler ve Çilo?lu, 1998).
Kideys ve ark. (1998), A?ustos 1993 döneminde 0-718 adet/m2 yumurtaya, 0-39 adet/m2 larvaya, Haziran-Temmuz 1996 döneminde ise 0-577 adet/m2 yumurtaya, 0-44 adet/m2 larvaya rastlam??lard?r. Niermann ve ark. (1994)’n?n A?ustos 1993’deki sörveyde bulduklar? ortalama yumurta say?s? (38.8 adet/m2), Temmuz 1992’deki sörveyde bulunan ortalama yumurta say?s?ndan (72.2 adet/m2) daha dü?üktür. Bununla birlikte ortalama en yüksek yumurta 89.5 adet/m2 de?eri ile Haziran-Temmuz 1996 sörveyinde
Türkiye’de toplam avc?l?k miktar? 1989-90 y?llar?nda 90000 ton’a (bu rakam?n içinde cinsi olgunlu?a eri?memi? bal?klar da vard?r) dü?mü?tür (Gucu, 2002). Hamsi miktar?nda görülen azalma 1992’den sonra avc?l?kta yasaklar?n gelmesi nedeniyle tekrar yükselmi?tir. Gucu (2002)’ye göre yeterli miktarda, h?zl? ve aktif olan küçük bal?k stoku ayn? besin kayna?? için ba?ar?l? rekabet içinde bulundu?u Mnemiopsis miktar?n? kontrol edebilecektir. Shiganova ve Bulgakova (2000) 1951 ve 1990 aral???ndaki çe?itli ara?t?r?c?lar?n mide içeri?i çal??malar?n? incelemi?ler özellikle hamside yetersiz besin içeri?ini 1989- 1990 aral???nda belirlemi?lerdir.
Kuzey Karadeniz’de Eylül (2002) ay?nda M. leidyi miktar olarak fazlaca bulundu?u dönemde zooplankton örnekleri 2001 y?l?ndan daha az bulunmu?tur. M. leidyi için ana besin kayna?? olan kladoser ve küçük kopepod miktarlar? oldukça dü?ük bulunmu?tur. 2001 y?l?nda s?cakl?k mevsim normallerinin üzerine ç?km?? ve bu dönemde B. ovata erken ortaya ç?km??t?r. Bu y?l?n A?ustos ay? ba??nda hamsinin yüksek yumurta ve larva de?erlerine ula??lm??t?r. B. ovata da??l?m?n?n geli?ti?i ilk iki y?l?n tersine larva miktar?nda yükselme gözlenmi?tir. 2002 y?l?nda ise yaz yumurtlama sürecinin bitti?i Eylül ay?nda toplanan yumurta 1999 ve 2001 y?l?ndakinden daha az say?da elde edilmi?tir (85 n/m2), fakat larva miktar? Beroe türünün ortaya ç?k???ndan önceki dönemlerden yüksek bulunmu?tur. Larvalar?n ço?unlu?unu hamsi (Engraulis encrasicolus ponticus), istavrit (Trachurus mediterraneus ponticus), barbunya (Mullus barbatus ponticus) ve isparoz (Diplodus annularis) türlerinin olu?turdu?u bildirilmi?tir (Shiganova ve Bulgakova, 2000; Shiganova ve ark., 2003; Shiganova ve ark., 2004a, 2004b). Toplam jelimsi organizma miktarlar? ve s?cakl?k aras?nda vertikal çekimlerdeki mevsimsel da??l?m ?ekil 5’te verilmi?tir.
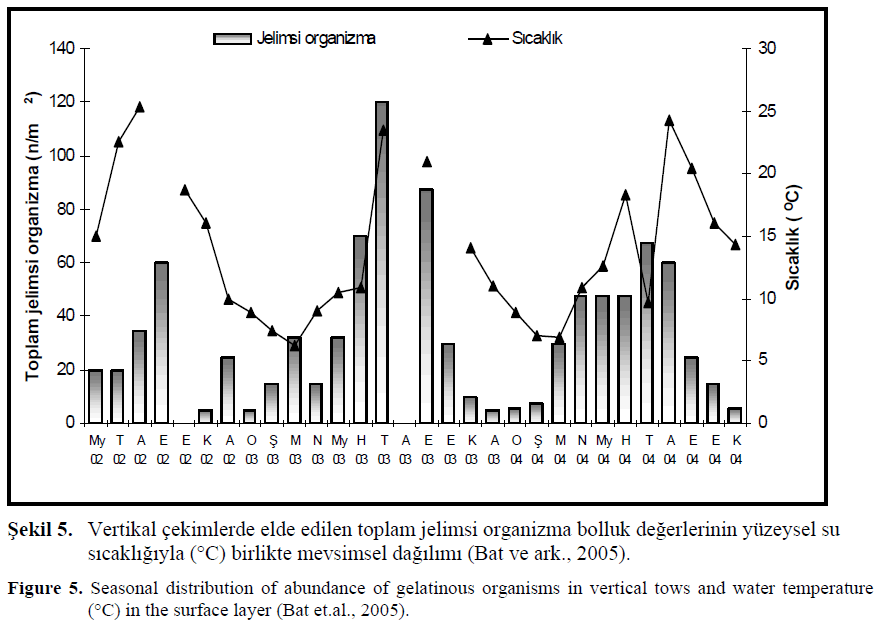
?ekil 5. Vertikal çekimlerde elde edilen toplam jelimsi organizma bolluk de?erlerinin yüzeysel su s?cakl???yla (°C) birlikte mevsimsel da??l?m? (Bat ve ark., 2005).
Figure 5. Seasonal distribution of abundance of gelatinous organisms in vertical tows and water temperature (°C) in the surface layer (Bat et.al., 2005).
Genel olarak bak?ld???nda s?cakl???n yükseldi?i ilkbahar, yaz ve sonbahar ba??nda toplam jelimsi organizma bolluk miktar?nda art?? gözlenmi?tir. Su s?cakl???n?n en yüksek ölçüldü?ü Temmuz 2003’de (23.5°C) toplam jelimsi organizmalar?n bolluk de?eri maksimum de?erine ula?m??t?r. Eylül ay?nda ise s?cakl?k de?eri 21°C oldu?unda da toplam jelimsilerin bolluk de?eri yüksek de?erlerde tespit edilmi?tir. 2004 y?l?nda yüzey suyu s?cakl???n?n Mart ay?ndan itibaren artmas? ile toplam jelimsi organizma miktar? da yükselmeye ba?lam?? ancak al?nan de?erler 2003 ile kar??la?t?r?ld???nda dü?ük oldu?u gözlenmi?tir. 2003 y?l?n?n Temmuz ay?nda yüksek bulunan jelimsi bolluk de?eri, 2004 y?l?n?n ayn? ay?nda daha çok dü?mü? ve bu ayda su s?cakl??? da oldukça (9.65 °C) azalm??t?r (Bat ve ark., 2005; Birinci-Özdemir, 2005). Sinop k?y?lar?nda ayn? zamana denk gelen deniz so?u?u olay?n?n bu farkl?l??a neden oldu?u dü?ünülmektedir. Ayr?ca bu ayda küçük boyda çok fazla miktarda M. leidyi bireyleri görülmü? fakat bireylerin oldukça dayan?ks?z oldu?u ve erime özelli?i gösterdi?i tespit edilmi?tir. Gözlem ve sonuçlara dayanarak s?cakl?k de?i?iminin canl?lar? morfolojik ve biyolojik aç?dan etkiledi?i dü?ünülmektedir.
Y?llar aras?nda yüzey suyu s?cakl??? de?i?imi paralellik göstermi?, ancak 2004 y?l?nda jelimsi miktar?n?n 2003 y?l?ndaki gibi maksimum de?erlere ula?mad??? görülmü?tür. Bununla beraber mevsimsel da??l?mda ilkbaharda bir önceki y?ldan daha fazla ve düzenli art???n oldu?u belirlenmi?tir.
Jelimsi organizmalar?n ana besinini zooplankton olu?turmaktad?r. Zooplankton miktar?n?n artmas? ile zooplankton üzerinden fazla miktarda beslenen jelimsi organizmalar?n bolluklar? artarken zooplankton miktar?nda dü?ü? meydana gelmektedir (Finenko ve ark., 2003, Shiganova ve ark., 2004a; 2004b).
Bu çal??mada A2 istasyonundaki vertikal çekimlerde zooplankton ve toplam jelimsi organizmalar?n bolluk de?erlerinin kar??la?t?r?lmas? ?ekil 6.’da verilmi?tir.
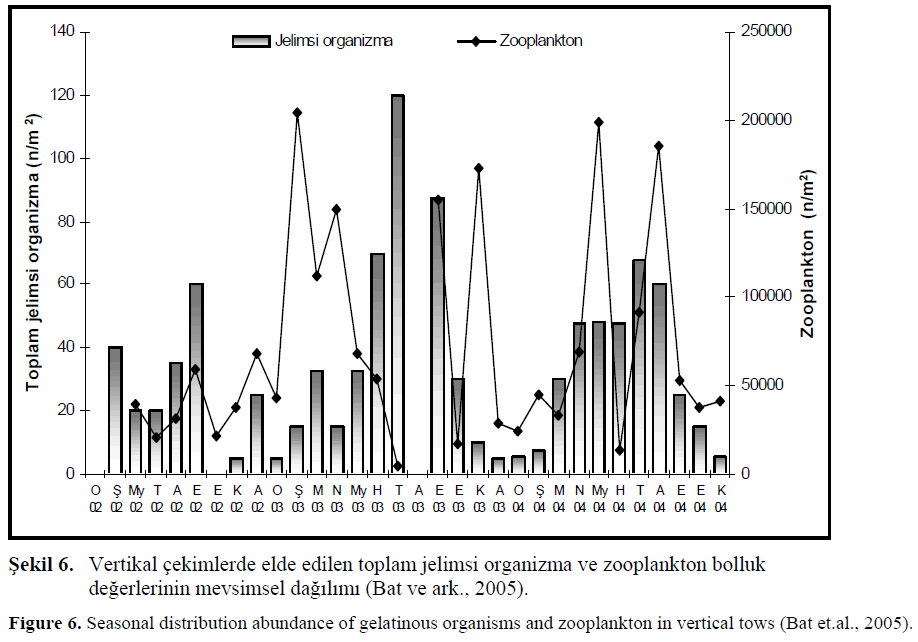
?ekil 6. Vertikal çekimlerde elde edilen toplam jelimsi organizma ve zooplankton bolluk de?erlerinin mevsimsel da??l?m? (Bat ve ark., 2005).
Figure 6. Seasonal distribution abundance of gelatinous organisms and zooplankton in vertical tows (Bat et.al., 2005).
2003 y?l?n?n k?? aylar?nda yüksek zooplankton miktar?na kar??l?k jelimsi organizma miktar?n?n dü?ük oldu?u, ilkbahar mevsiminde jelimsi organizma miktar?n?n artmaya ba?lamas? ile zooplankton miktar?n?n azald??? gözlenmi?tir. ?ubat ay?nda zooplankton bolluk de?eri maksimum 200 000 n/m2’ye ula?t???nda jelimsi organizma bolluk miktar?n?n 15 n/m2 dü?tü?ü, Temmuz ay?nda ise bunun tersine jelimsi organizma maksimum bolluk de?erini al?rken zooplankton miktar? 4800 adet/m2 dü?mü?tür. Eylül ay?nda zooplankton bolluk de?eri artm?? jelimsi organizma bolluk de?eri bir miktar dü?mü?, Ekim ay?nda her iki grubun bolluk de?erlerinin dü?tü?ü tespit edilmi?tir. Bu aylar? takip eden di?er aylarda ise tekrar zooplanktonun yüksek de?ere ula?t??? ve jelimsi organizma miktar?n?n dü?tü?ü görülmü?tür. Biyokütle miktarlar?nda ise söz konusu aylarda dü?ük de?erler al?nm??t?r. Bu aylarda P. pileus, M. leidyi ve B. ovata türlerinin say?ca fazla ancak a??rl?klar?n?n az olmas?, biyokütle de?erlerinin dü?ük ç?kmas?na yol açm??t?r (BirinciÖzdemir, 2005).
2004 y?l?nda zooplankton miktar? k?? aylar?nda dü?ük seviyede kalm?? ancak ilkbahar ba??nda artmaya ba?lam??t?r. May?s, Temmuz ve A?ustos aylar?nda zooplankton bolluk ve biyokütle miktarlar?nda yüksek de?erlere ula??lm??t?r. Bir önceki y?l (2003) ile kar??la?t?r?ld???nda genel olarak jelimsi organizmalar?n yo?un oldu?u aylarda zooplankton miktar?nda art?? belirlenirken, jelimsi organizma miktar?nda ise azalma görülmü?tür. Özellikle 2004 y?l?nda zooplankton ve jelimsi organizmalar?n biyokütle de?erlerindeki de?i?imler, zooplankton miktar?ndaki iyile?menin önceki y?llar ile kar??la?t?r?lmas?na ve daha iyi yorumlamas?na imkan vermi?tir.
Yumurta ve larva bolluk miktar? ile jelimsi organizma ve zooplankton miktarlar?n?n vertikal çekimlerdeki bolluk de?erleri ?ekil 7.’de verilmi?tir. Genel olarak bak?ld???nda bal?k yumurta ve larvalar?n?n artt??? dönemlerde jelimsi organizma miktar?nda art?? kaydedilmi?tir. Örne?in 2002 y?l?n?n Aral?k ay?nda, 2003 May?s ay?ndan ba?layarak Ekim ay?na kadar, 2004 Mart ay?ndan ba?layarak ve yine Ekim ay?na kadar süren bir art?? gözlenmi?tir. Ayl?k olarak incelendi?inde her bir ay için jelimsi ve bal?k yumurta ve larvas? aras?nda tam olarak do?rusal bir paralellik bulunamamas?na ra?men y?ll?k de?i?imler incelendi?inde ili?ki daha net tespit edilmi?tir. 2004 y?l?nda di?er iki örnekleme y?llar?na oranla yumurta ve özellikle larva miktar?nda art?? belirlenirken jelimsi miktar?nda azalma dikkat çekmi?tir. Hamsi üreme dönemi olan yaz mevsiminde 2004 y?l? incelendi?inde zooplankton bolluk ve özellikle biyokütle miktar?n?n artmas? ile bal?k yumurta ve larva miktar?nda art?? belirlenmi?tir. Bunun yan?nda jelimsi organizma miktar?n?n azalmas? yumurta ve larva miktarlar?n?n artmas?nda etkili oldu?u tahmin edilmektedir (Bat ve ark., 2005; Birinci-Özdemir, 2005).
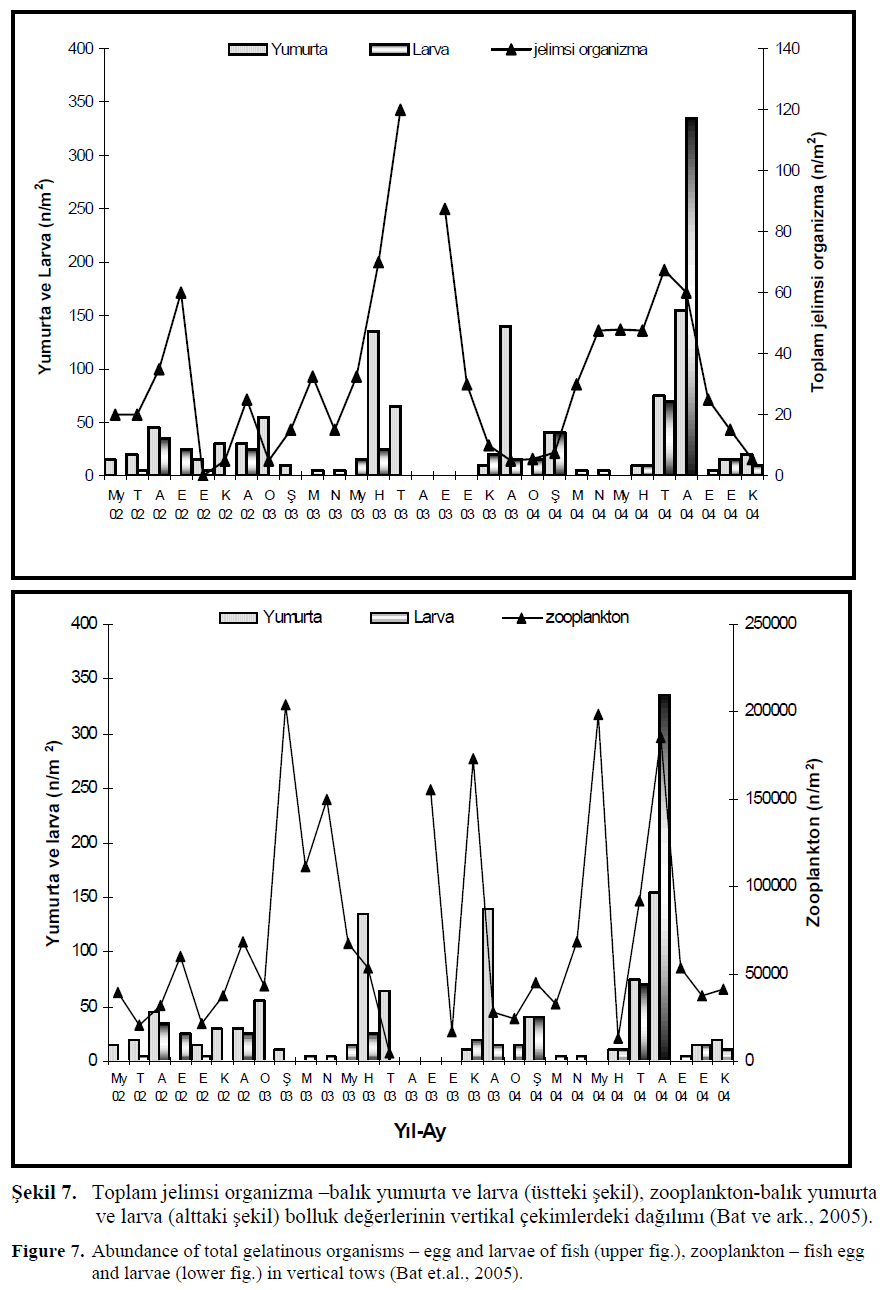
?ekil 7. Toplam jelimsi organizma –bal?k yumurta ve larva (üstteki ?ekil), zooplankton-bal?k yumurta ve larva (alttaki ?ekil) bolluk de?erlerinin vertikal çekimlerdeki da??l?m? (Bat ve ark., 2005).
Figure 7. Abundance of total gelatinous organisms – egg and larvae of fish (upper fig.), zooplankton – fish egg and larvae (lower fig.) in vertical tows (Bat et.al., 2005).
1980’li y?llarda ötrofikasyon, a??r? avc?l?k ve M. leidyi türünün Karadeniz’e giri?iyle bal?k avc?l???nda ve özellikle ekonomik de?ere sahip hamsi miktar?nda tekrar azalma ba?lam??t?r (?ekil 8., Caddy ve Griffiths, 1990; Kideys ve ark., 1999; Shiganova ve Bulgakova, 2000; Kideys, 2002).
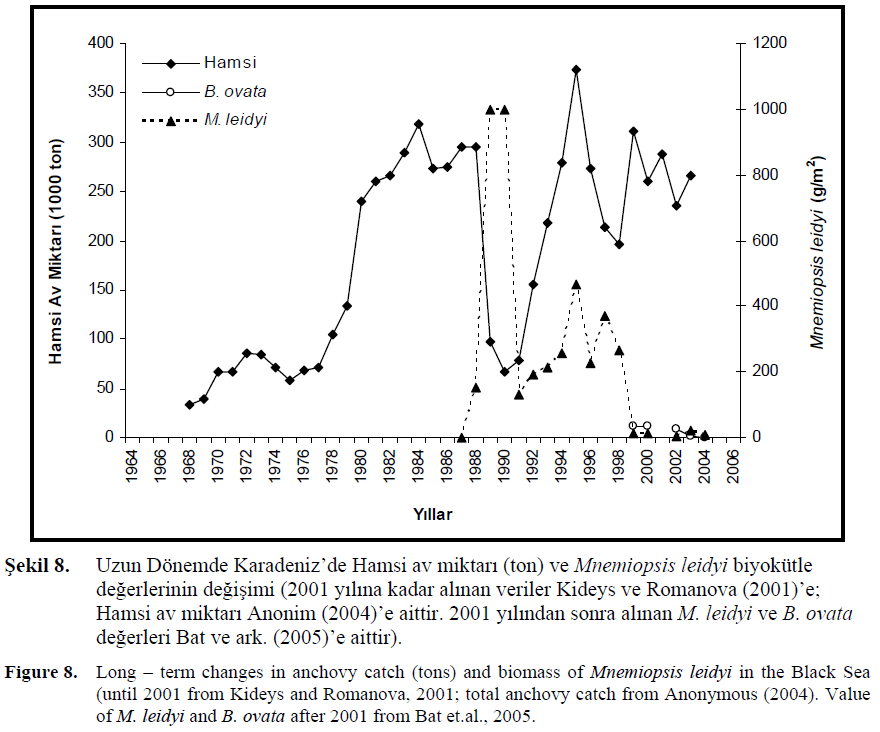
?ekil 8. Uzun Dönemde Karadeniz’de Hamsi av miktar? (ton) ve Mnemiopsis leidyi biyokütle de?erlerinin de?i?imi (2001 y?l?na kadar al?nan veriler Kideys ve Romanova (2001)’e; Hamsi av miktar? Anonim (2004)’e aittir. 2001 y?l?ndan sonra al?nan M. leidyi ve B. ovata de?erleri Bat ve ark. (2005)’e aittir).
Figure 8. Long – term changes in anchovy catch (tons) and biomass of Mnemiopsis leidyi in the Black Sea (until 2001 from Kideys and Romanova, 2001; total anchovy catch from Anonymous (2004). Value of M. leidyi and B. ovata after 2001 from Bat et.al., 2005.
Hamsi av miktar?nda 1980’li y?llar?n sonunda 5 kattan fazla bir azalma belirlenmi?tir (Kideys, 1994; Gucu, 1997). Karadeniz’de hamsi av miktar? 62 bin ton seviyesine dü?mü?, ayn? dönemde M. leidyi biyokütle miktar? 1000 g/m2 hesaplanm??t?r. Bu dü?ü?ün ard?ndan 90’l? y?llar?n sonunda yine stabil ?ekilde art?? ba?lam?? ve hamsi av miktar? 390 bin ton seviyesine yükselmi?tir, bu de?er M. leidyi Karadeniz’e geldikten sonra al?nan en iyi miktard?r. Yine ayn? dönemde M. leidyi biyokütle de?eri incelenmi? ve 500 g/m2 de?eri bulunmu?tur. Bu M. leidyi miktar?ndaki azal???n bir göstergesi olmu?tur. Uzun dönemdeki M. leidyi biyokütle ve Karadeniz hamsisi miktar? aras?ndaki de?i?imlere paralel olarak z?t bir ili?kinin oldu?u söylenmektedir (Kideys ve Romanova, 2001; Shiganova ve ark., 2003, ?ekil 6.5.4.). M. leidyi türünün neden oldu?u de?i?imler sadece miktarlarda olmam?? daha önceden Karadeniz’in Kuzeybat? bölümü hamsi için yumurtlama alan? iken Güney Karadeniz’in yumurtlama bölgesi olarak daha yo?unla?t??? Niermann ve ark. (1994) taraf?ndan belirtilmi?tir.
M. leidyi türünün mevsimsel büyümeleri yaz ve sonbahar ba??nda hamsi ve di?er planktonla beslenen bal?klar?n üreme mevsimiyle ayn? döneme denk gelmektedir. Mnemiopsis bal?k larvalar?n?n onda birini avlayarak veya besin içeri?inde kullanarak miktarlar?n? de?i?tirmi?lerdir. Niermann ve ark. (1994) taraf?ndan Mnemiopsis ve Karadeniz hamsisi (E. e. ponticus) aras?ndaki ili?ki 1980’lerin sonu ve 1990’lar?n ba??nda rapor edilmi?tir.
Karadeniz’de hamsi av miktar?ndaki azal???n nedenleri 4 hipotez ile özetlenebilir (Bilio ve Niermann, 2004).
Birinci hipotez; M. leidyi türünün a??r? miktarda bal?k yumurta ve larvas?n? tüketmesidir. 1988 ile kar??la?t?r?ld???nda K?r?m Yar?madas?’nda bal?k yumurta ve larva miktar?n?n azald??? dikkat çekmi?tir (Niermann ve ark., 1994). M. leidyi türünün bal?k yumurta ve larvas?n?n predatörü oldu?u saptanm?? olmas?na ra?men (Cowan ve Houde, 1993), tüm azalman?n bu türden kaynakland??? kesin de?ildir. Tsikhon-Lukanina ve ark. (1991 ve 1993) taraf?ndan M. leidyi türünün kopepod üzerinde bal?k yumurta ve larvas?ndan daha yo?un olarak beslendi?ini tespit edilmi?tir. Bal?k yumurta ve larvas?n?n ani dü?ü?üne ek olarak M. leidyi bolluk miktar? yüksek seviyeye ula?m?? ve bu art??, bal?k miktar?ndaki azalmay? beraberinde getirmi?tir. Dolay?s?yla M. leidyi türünün bu azalmadaki etkisi önemli bir faktör olarak göz önünde bulundurulmal?d?r (Bilio ve Niermann, 2004; Shiganova ve ark., 2004a; 2004b).
?kinci hipotez M. leidyi türünün a??r? miktarda zooplankton tüketmesi ile hamsi stoklar?ndaki dü?ü? olarak tan?mlanmaktad?r. Karadeniz geneline bak?ld???nda 1989-1992 y?l?na Mnemiopsis bolluk miktar?n?n oldukça artmas? ile ayn? zamana gelen mesozooplankton miktar?ndaki dü?ü?ten dolay? bu hipotez daha makul gelmektedir (Konsulov ve Kamburska, 1998; Kovalev ve ark., 1998a; Kideys ve ark, 2000). Mezozooplankton miktar?n?n azalmas?yla birlikte plankton ile beslenen di?er bal?k stoklar?n?n (Çaça) besin sorunu çekti?i görülmü?tür (Shulman ve Yuneva, 2002).
Kideys ve ark. (2000) hamsinin ana besininde belirlenen kötüye gidi?le beraber a??r? ve önemli tüketici M. leidyi 1989-1990 y?llar? aras?nda bal?k stoklar?nda ?iddetli bir dü?ü?e yol açt??? belirtmi?lerdir. ?lk önce M. leidyi türünün önce kopepod Calanus euxinus nauplileri ile beslenerek hamsi larvalar?n? etkiledi?ini daha sonra yeti?kin hamsi için daha ilerideki ergin kopepod büyüklü?ünü seçerek beslendi?ini bu yüzden bal?k avc?l???nda 1990’larda dü?ü?ün meydana geldi?ini varsaym??lard?r.
M. leidyi miktar?ndaki art?? ile hamside meydana gelen dü?ü? o kadar kesi?mektedir ki bu iki olay?n do?rudan do?ruya birbiriyle ba?lant?l? oldu?u san?lmaktad?r. Olaylar?n ayn? zamana rastlamas? üzerinde özellikle fazla durulmas?n? önemli hale getirmektedir. Vinagradov ve ark. (1989) taraf?ndan jelimsi zooplankton da??l?m?n?n bölgesel da??l?m?n?n belirlenmesinde 32 istasyonda örnekleme yap?lm?? ve biyokütle de?eri yakla??k 84.107 g olarak Karadeniz’de tespit edilmi? ve istilac? ktenofor türünün mevcut mesozooplankton ve bal?k larvalar? stoklar?n? ciddi ?ekilde etkiledi?i gösterilmi?tir.
A??r? avc?l?k, hamsi stoklar?n?n azalmas?ndaki üçüncü hipotezi olu?turmaktad?r. 1980’lerin sonunda Karadeniz’de bal?kç?l?k çe?itli nedenlerle karma??k hale gelmi?tir. Sovyetler Birli?i’nde bal?kç?l?k ekonomik sebepler ba?ta olmak üzere azalm?? bu dönemde Türk bal?kç?lar? teknelerinin bal?k bulucu cihazlar?n? ve güverte üstü donan?mlar?n? geli?tirmi?lerdir (Gucu, 2002). Bu nedenle Türk bal?kç?l???ndaki yükseli?le birlikte Sovyetler Birli?i avc?l?k alan?nda dü?ü? ya?am??t?r (Bilio ve Niermann, 2004).
Gucu (1997), 1988-1989 y?llar?ndaki Türkiye k?y?lar?ndan al?nan verileri 1987-1988 verileri ile kar??la?t?rd???nda bal?k boylar?n?n daha küçük boyda olduklar?n? rapor etmi?tir. Hamsi stoklar?nda 1988-1989 meydana gelen dü?ü?, bal?kç? teknelerinin geli?imi ile önceki y?llardaki av miktar?n? yakalamaya çal???rken cinsel olgunlu?a ula?mam?? bal?klar?n ve küçük boydaki bal?klar?n avlanmas? neticesinde olu?tu?u dü?ünülmektedir.
Türkiye sular?ndaki bal?kç?l?k endüstrisinin çökü?ünden önce ve sonra yap?lan hamsi gonadosomatik indeks analizleri, hamsi stoklar?nda meydana gelen çökü?ten sonra yeni kat?lan bireylerin durumunun çökme öncesine göre daha iyi oldu?unu göstermi?tir. Hamsi stoklar?n?n tüketilmesi ile birlikte var olan besin miktar?n?n ve di?er planktivor organizmalar?n artmas? sürpriz olmam??t?r (Bilio ve Niermann, 2004). Bal?k avc?l??? miktar?nda ani dü?ü?ün ana sebebi 1988’den önce bal?kç?l?k ekipmanlar?n?n geli?mesiyle a??r? avc?l???n artmas? olarak görülmektedir (Gucu, 1997). Uzun dönemde mide ve barsak içeriklerinin incelendi?i veriler (Shiganova ve Bulgakova, 2000) özellikle hamsinin, bal?k stoklar?n?n azald??? döneme kadar yeterli besine sahip oldu?unu göstermi?tir.
Dördüncü hipotez ise o?inografik ve meteorolojik do?a olaylar?n?n etkisiyle sistemin de?i?im ya?amas?d?r. 1980’lerde meydana gelen hidrolojik ve meteorolojik olaylar Kuzey Atlantik’ten Karadeniz ve Hazar Denizi’ne do?ru kuzey yar?mkürede de?i?ime neden olmu?tur (Niermann ve ark., 1999; Oguz, 2005b).
Genel olarak özetlenirse Karadeniz’de ekosistem 2 ?ekilde etkilenmi?tir; (1) a?a??dan yukar?ya etki; 1970’li y?llarda nehirler ile k?y? sahil sular?n?n dip k?sm?na ula?arak içerideki termoklin tabakas?na kadar uzanan nutrientlerin olu?turdu?u aktif organik madde sirkülasyonu ve sonras?nda antropojenik güçler, (2) yukar?dan a?a??ya etki ise; pelajik bal?klar?n a??r? avc?l??? ve 1980’li y?llarda hüküm süren karnivor jelimsi organizmalar?n a??r? populasyon art???d?r. Bu e? zamanl? etkiler y?llarca al?nan datalar?n birle?tirilmesi ile ortaya ç?kar?lm??t?r (O?uz, 2005a).
Nutrientlerin Karadeniz’de artmas? ile birlikte ötrofikasyonun etkisi, mesozooplankton yap?s? ve da??l?m?nda, bal?k stoklar?n?n azalmas?nda ve a??r? alg art??lar?nda gözlenmeye ba?lanm??t?r. Ekosistem dengesinin bozulmaya ba?lamas?yla jelimsi organizmalarda de?i?imler olmu?tur. ?öyleki; Karadeniz’de a??r? avc?l?k ile büyük bal?k miktar? azalm??, küçük pelajik bal?klar ise pelajik bolluk için predatör olmu?lard?r. Mesozooplankton miktar?n?n de?i?imiyle fitoplankton de?erleri yükselmi?tir (Shiganova, 1998; Gucu, 2002; Daskalov, 2002). 1980’lerin sonunda jelimsi organizmalar sistemi kontrol alt?na ald???nda ve küçük pelajik bal?k stoklar? tüketildi?inde a??r? avc?l???n tahripleri fark edilmi?tir.
Daha öncede belirtti?imiz gibi ötrofikasyon ve a??r? avc?l?k ekosistemi özellikle 1990’l? y?llarda çok güçlü olarak etkilemi? görülmemektedir (O?uz, 2005a). Bunun yan?nda M. leidyi türünün a??r? miktarda artmas?, sistemi etkileyen di?er bir unsur olmu?tur. 1990’l? y?llarda M. leidyi biyokütlesi bir miktar dü?mü?, bal?k avc?l??? 1992 ve 1993 y?llar?nda minimum de?erini alm??t?r. 1995 y?l?nda bal?k avc?l??? yükselmi? ve yine 1998 y?l?nda tekrar dü?mü?tür. 1999’dan ba?layarak ktenofor B. ovata, M. leidyi türünün tüketilmesinde ve ekosistemin iyile?mesinde etkili olmu?tur. Ayn? zamanda tekrar bal?k avc?l??? yükselmeye ba?lam??t?r (Kideys ve ark., 1999; Kideys ve ark., 2000; Kideys ve Romanova, 2001; Anonim, 2004; Shiganova ve ark., 2004a; 2004b).
Finenko ve ark. (2003) 2000-2001 y?llar? ile (1.10±0.65 g/m2) 1995 y?l?n? (0.42±0.38 g/m2) kar??la?t?rd?klar?nda, ortalama y?ll?k zooplankton biyokütlesinin iki kat artm?? oldu?unu tespit etmi?lerdir. Bu farkl?l???n istatistiksel olarak önemli oldu?u bulunmu?tur (p<0.05). Mnemiopsis türünde ani art??lar?n oldu?u ve Beroe türünün bulunmad??? önceki y?llar ile kar??la?t?r?ld???nda, 2000-2001 y?llar?nda baz? türlerin (P. parvus, A. clausi, C. ponticus) ve özellikle ortalama y?ll?k kopepod bolluk miktar?n?n yükseldi?i bildirilmektedir.
Bu çal??mada jelimsi organizmalardan M. leidyi ve B. ovata türlerinin bolluk de?erleri zooplankton bolluk ve biyokütle de?erleri ile kar??la?t?r?lm??t?r. M. leidyi bolluk art???n? takip eden bir sonraki ayda zooplankton miktar?n?n azald???, B. ovata türünde ise tam tersi zooplankton miktar?n?n art?? gösterdi?i tespit edilmi?tir. Bununla birlikte M. leidyi miktar?n?n dü?ük oldu?u dönemlerde az miktarda besin tüketilmesinden dolay? zooplankton bolluk ve biyokütle miktar?n?n yükseldi?i dü?ünülmektedir. Temmuz (2003) ay?nda M. leidyi bolluk ve biyokütle de?eri maksimum de?erlere ula?t???nda, zooplankton bolluk ve biyokütle miktar?n?n örneklemelerdeki en dü?ük de?erini alm??t?r. Sonraki A?ustos ay?nda ise M. leidyi miktar?n?n dü?mesi ile zooplankton bollu?unun artt??? belirlenmi?tir (?ekil 9).
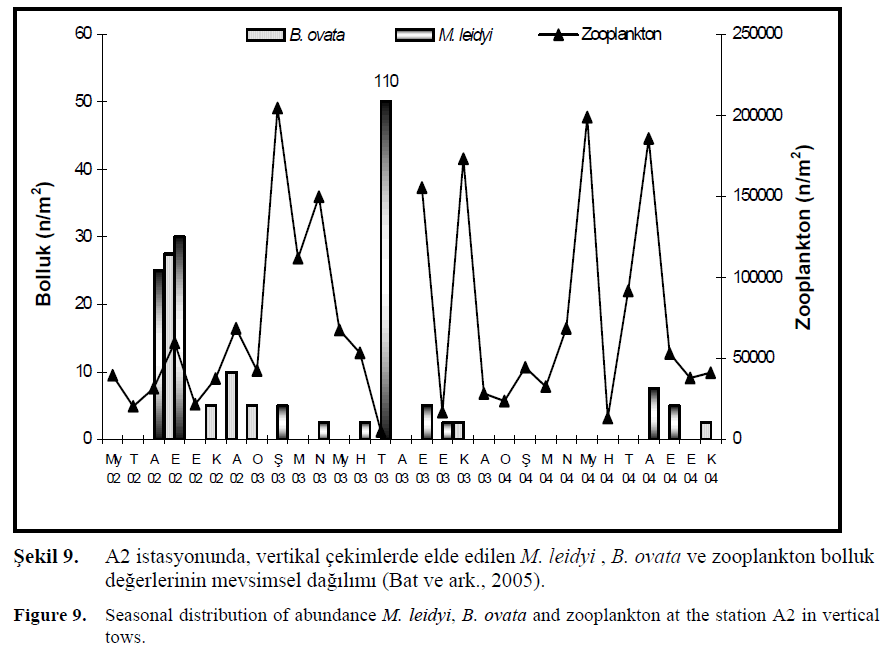
?ekil 9. A2 istasyonunda, vertikal çekimlerde elde edilen M. leidyi , B. ovata ve zooplankton bolluk de?erlerinin mevsimsel da??l?m? (Bat ve ark., 2005).
Figure 9. Seasonal distribution of abundance M. leidyi, B. ovata and zooplankton at the station A2 in vertical tows.
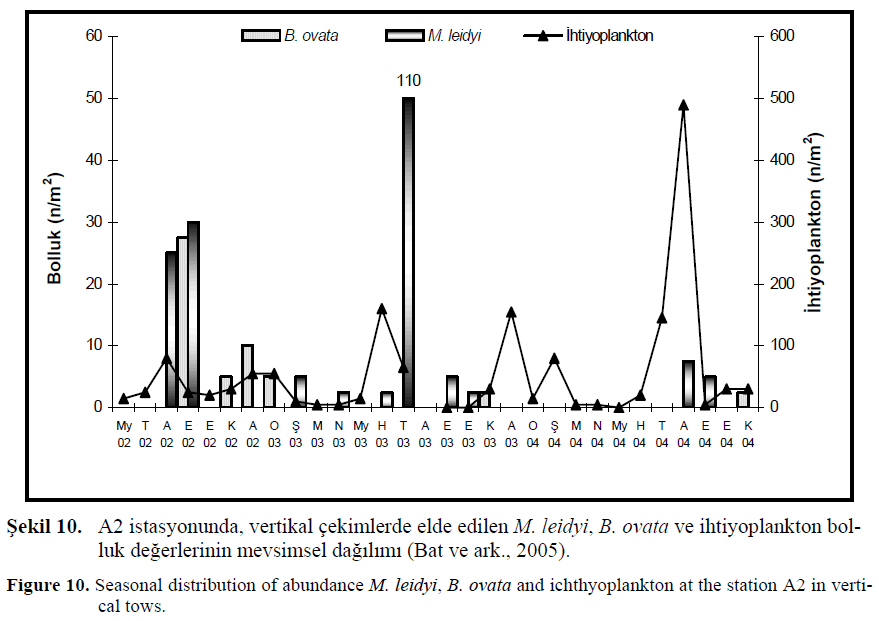
?ekil 10. A2 istasyonunda, vertikal çekimlerde elde edilen M. leidyi, B. ovata ve ihtiyoplankton bolluk de?erlerinin mevsimsel da??l?m? (Bat ve ark., 2005).
Figure 10. Seasonal distribution of abundance M. leidyi, B. ovata and ichthyoplankton at the station A2 in vertical tows.
Bat ve ark., (2005) taraf?ndan yap?lan çal??mada ihtiyoplankton bolluk miktar? ile M. leidyi ve B. ovata bolluk de?erlerini aras?nda aç?k bir ili?ki bulunamam??t?r. A?ustos (2004) ay?nda ihtiyoplankton miktar?nda ani bir art?? belirlenmi?tir. ?öyle ki Karadeniz’de bal?k miktar?n?n büyük bölümünü (% 85) hamsi olu?turmaktad?r (Anonim, 2004). Hamsisi’nin üreme zaman? yaz dönemi oldu?undan böyle bir art???n olmas? beklenen bir durumdur. Fakat üç y?l süre ile yap?lan örneklemelerde önceki iki y?l?n hamsi üreme dönemlerinde al?nan veriler 2004 y?l? A?ustos ay?ndan oldukça dü?üktür.
Sonuç olarak Sinop Yar?madas?’nda yap?lan mevcut çal??mada s?cakl?k de?erlerinin yükseldi?i dönemlerde jelimsi organizma miktar?n?n artt??? tespit edilmi?tir. S?cakl???n artmaya ba?lad??? ilkbahar mevsiminde jelimsi bolluk ve biyokütle düzeyinde art??lar belirlenmi?tir. Yaz mevsiminde maksimum de?erlere ula?an jelimsi organizma miktar? sonbahar mevsimi sonuna kadar azalarak örneklemelerde gözlenmeye devam etmi?tir. Türlerin üreme dönemlerinde bolluk ve biyokütle miktarlar?nda görülen art??lar beklenen bir sonuçtur. K?sa süreli ayl?k incelemelerde zooplankton ve ihtiyoplankton ile jelimsi organizmalar?n miktarlar? aras?nda bir paralellik bulunamam?? olup, üç y?ll?k uzun dönemdeki örnekleme sürecine bak?ld???nda paralellik aç?kça görülmü?tür. Bunun yan?nda jelimsi organizma miktar?n?n art??? ile ihtiyoplankton miktar? aras?ndaki ba?lant?n?n zooplankton ile olan ba?lant?s?ndan daha dü?ük oldu?u tespit edilmi?tir. Çal??mada elde edilen bu bilgiler di?er çal??malar ile ayn? paralellikte belirlenmi?tir. Özellikle M. leidyi türü ile zooplankton biyokütle ve bolluk de?erleri aras?ndaki ters orant? bu çal??mada bir kez daha ortaya konulmu?tur. Bu çal??ma süresince 2004 y?l?nda al?nan jelimsi organizmalar?n miktar?ndaki dü?ü? ile ihtiyoplankton, zooplankton miktar?ndaki art??lar ve hamsi avc?l???ndaki art???n sistemdeki iyile?melerin ve düzelmelerin bir göstergesi oldu?u dü?ünülmektedir.
Tüm bunlarla beraber Karadeniz’deki de?i?imler ve ekosisteme etkileri oldukça önemlidir. Sistemde meydana gelen de?i?imler yap?y? kökten etkilemekte ve düzelmesi imkans?z veya çok zor olan zararlar? beraberinde getirmektedir. Bu nedenle Karadeniz ekosisteminde 1980’lerden sonra bask?n duruma geçen jelimsi organizmalar? tan?mak, etkilerini ve da??l?mlar?n? incelemek gerekli hale gelmi?tir. Özellikle hamsi avc?l???n?n ekonomik aç?dan yüksek de?er ta??mas? çal??malar? bir kat daha önemli k?lm??t?r. Karadeniz’e k?y?s? olan baz? ülkelerde jelimsi organizmalar ile ilgili çal??malar devam ettirilmektedir. Kuzey Karadeniz’de yap?lan çal??malarla sistematik olarak veriler al?nmaktad?r. Özellikle Karadeniz’in Ülkemiz k?y?lar?nda uzun dönemi kapsayacak zaman serili çal??malar bulunmamaktad?r. Ancak son 6 y?ld?r Sinop Su Ürünleri Fakültesi Temel Bilimleri Bölümünce sürdürülen TÜB?TAK, DPT ve NATO destekli projelerde uzun periyotlu çal??malar devam etmektedir (Büyükhatipo?lu ve ark., 2002; Bat ve ark., 2004a,b ve 2005; Bircan ve ark., 2005). De?i?imlerin incelenmesi, çözüm önerilerinin sa?lam bir zeminde yürütülmesi aç?s?ndan sistemdeki fiziko-kimyasal parametrelerin, zooplankton, fitoplankton, ihtiyoplankton ve jelimsi organizmalar?n da??l?mlar?n?n (de?i?imlerinin) izlenmesinin oldukça önemli olmas? yürütülen çal??malar?n devaml?l???n? zorunlu k?lmaktad?r. Ayr?ca bal?k türleri ve stok hareketleri ile ilgili ara?t?rmalarla bu çal??malar?n desteklenmesi yararl? olacakt?r.
Karadeniz kendisine has yap?s?yla h?zl? fiziksel ve kimyasal de?i?imin yan?nda özellikle istilac? türler bak?m?ndan h?zla de?i?ebilen ve yeni dengelere kavu?abilen özel bir denizdir. Ülkemiz bal?kç?l???ndaki önemini de dikkate al?rsak hem denizin özelikleri hem de ya?ayan organizmalar ve bal?klar üzerinde her y?l düzenli ve detayl? çal??malar?n sürdürülmesi gerekmektedir. Dolay?s?yla yo?un i? gücü ve yüksek bütçeyi gerektiren ve de günümüze de?in eksikli?i büyük oranda hissedilen deniz ara?t?rmalar? istenen seviyeye ula?acakt?r.
Uzun süreli gözlemlere dayal? verilerin önemi bilhassa olas? iklim de?i?ikliklerinin analizi aç?s?ndan günümüzde büyük bir öneme sahiptir. ?klim de?i?ikliklerinin yan?nda günümüzde her geçen gün artan kirlili?in ekosistem üzerine olan etkilerinin kalitatif ve kantitatif olarak ortaya konmas? için uzun süreli gözlemlere ihtiyaç vard?r. Karadeniz’i çevreleyen baz? ülkelerin Karadeniz’de izleme çal??malar? 1950’lere kadar uzan?rsa da 1980’li y?llarda demirperde olarak adland?r?lan bu ülkelerde ekonomik durumun bozulmas?yla 1980’lerden sonra kesintiye u?ram??, bu tarihten sonraki veriler bir ya da birkaç istasyonda k?sa aral?kl? uzun döneme ait veri elde edilmesinden çok y?lda bir-iki kez gerçekle?tirilebilen seferlerden elde edilmi?tir. Bu verilerin analizi hem fitoplankton ve hem de zooplanktonda önemli de?i?imlerin oldu?una i?aret etmektedir. Denizlerde biyojeokimyasal döngülerin anla??lmas?nda için tüm denizden veri elde etmeyi sa?layan kapsaml? seferlerin yan?nda, bir ya da birkaç istasyondan k?sa aral?kl? veri toplamak gerekmektedir. Ancak böyle bir veri seti elde ettikten sonra o denizde anahtar öneme sahip canl?lar?n temel (üreme, büyüme, ölüm vs gibi) dinamikleri anla??labilir. Karadeniz’de planktonun mevsimsel da??l?m?n? tüm y?l boyunca düzenli aral?klarla veren çal??malar azd?r. Mevcut çal??mada ise, Orta Karadeniz’de temel pelajik ekosistem parametrelerinin izlenmesi çerçevesinde, hem besleyici elementlerin hem de bu besleyici elementlerden faydalanan fitoplankton, zooplankton, ihtiyoplankton ve makrozooplanktonun mevsimsel yap?s?n?n tan?mlanmas? ve uzun dönem de?i?ikliklerin izlenmesi amaçlanm??t?r.
Sonuç olarak bu çal??ma, bundan sonra yap?lmas? planlanan ekosistem modellemelerine veri taban? olu?turma ve ekosistemin rehabilitasyonu yönünde karar al?c? mercilere done olu?turma aç?s?ndan irdelendi?inde kapsam ve katk?lar? aç?s?ndan büyük önem ta??yacakt?r.
Te?ekkür: Bu çal??ma, NATO (NATO ESP. NUKR. CLG. 981783) nolu “Bioindicators for Assessment of the Black Sea Ecosystem Recovery” ve TÜB?TAK-NASU 105Y028 nolu “Monitoring of anchovy and sprat nutritional condition in the Black Sea” projeler taraf?ndan desteklenmi?tir.
733
References
- Anonim, (1968-1990). DIE (Devlet Istatistik Enstitüsü), Su ürünleri 1968-1990 Verileri, Ankara Anonim, (1997). Opportunistic Settlers and the Problem of the Ctenophore Mnemiopsis leidyi Invasion in the Black Sea. Gesamp Reports and Studies no. 58. Printed by the International Maritime Organization, 4 Albert Embankment, London SE1 7SR
- nAnonim, (2004). (DIE) Devlet Istatistik Enstitüsü, Su ürünleri 2003 Verileri, Ankara
- nArim, N., (1957). Marmara ve Karadeniz’deki Bazi Kemikli Baliklarin (Teleostlarin) Yumurta ve Larvalarinin Morfolojileri ile Ekolojileri. Hidrobiyoloji Mecmuasi, Cilt 5(1-2): 7-55
- nBat, L., Kideys, E.A., Oguz, T., Besiktepe, S., Yardim, Ö., Gündogdu, A., Üstün, F., Satilmis, H.H., Sahin, F., Birinci Ö.Z. ve Zoral, T., (2005). Orta Karadeniz’de temel pelajik ekosistem parametrelerinin izlenmesi. Proje no: DPT 2002 KI20500 (TAPS013 No’lu Proje). 488 s
- nBat, L., Kideys, A., Yardim, Ö., Gündogdu, A., Üstün, F., Satilmis, H.H., Sahin F., Birinci Ö.Z. ve Zoral, T. (2004a). Sinop Kiyilarinda Fitoplankton, Zooplankton, Ihtiyoplankton ve Makrozooplankton Kompozisyonunun Karadeniz Ekosistemine ve Balikçiliga Etkileri. O.M.Ü. S.090 Nolu Arastirma Fonu Projesi. 130 s
- nBat, L., Finenko, G.A., Anninsky, B.E., Romanova, Z.A., Svetlichny, L.S., Abolmasova, G.I., Hubareva E.S. and Kideys, A., (2004b). Will the new Invader Ctenophore Beroe ovata Control the Structure of Plankton Community in the Black Sea? (NATO Scientific Affairs Linkage Grant EST. CLG. 976805
- nBayraktar, S., 1994. Distrubition of phytoplankton (>55 um) along Turkish Coast and at The North Western Shelf area of The Black Sea (Yüksek Lisans Tezi), ODTÜ, 172 s
- nBenli, H., (1987). Investigations of plankton distribution in the southern black Sea and its effects on particle flux. In: Degens, E. T., Izdar, E., Honjo, S. (eds) Particle Flux in the Ocean, Mitteilungen des Geologisch- Paleontologischen Institut, Universitat Hamburg, 62: 77-87
- nBerdnikov, S. V., Selyutin, V., V., Vasilchenko, V. V. and Caddy, J.F., (1999). Trophodynamic model of the Black and Azov Sea pelagic ecosystem: consequences of the comb jelly, Mnemiopsis leidyi, invasion. Fisheries Research, 42:261-289
- nBesiktepe S., (2001). Diel vertical distribution, and herbivory of copepods in the south western part of the Black Sea. Journal of Marine Systems, 28: 281-301.
- nBingel, F., Kideys, A. E., Özsoy, E., Tugrul, S., Bastürk, Ö. and Oguz, T. (1993). Stock Assessment Studies For The Turkish Black Sea Coast. NATO-TU Fisheries Final Report. METU, Institute Of Marine Sciensec,
- nBircan, R., Bat, L., Kideys, A., Satilmis, H.H., Üstün, F., Sahin, F. ve Özdemir, Z. B., (2005). Karadeniz’in Sinop Bölgesinin Alt Besin Tabakalarinin Dinamik Ve Zaman Serileri, Tübitak, Proje No: 199Y121, 75s
- nBirinci-özdemir, Z., (2005). Karadeniz’in Sinop Burnu Bölgesinin Jelimsi Organizma Kompozisyonu ve Mevsimsel Dagilimi. Fen Bilimsleri Enstitüsü Su Ürünleri Temel Bilimleri Anabilim Dali, Yüksek Lisans Tezi, 147 s
- nSamsun. Bodeanu, N., (1989). Algal blooms and development of the main phytoplanktonic species at the Romanian Black Sea littoral under eutrophication conditions. Cercet. Mar. I. R. C. M. 22:107-125
- nBologa, A. S., (1985/1986). Planktonic primary productivity of the Black Sea: review. J. Thalassia Jugoslavica, 21/22 (1/2): 1-22
- nBrink, K.H., (1978). Upwelling Fronts; Implications and Unknows. S.A., Sc., 5: 3- 9
- nBüyükhatipoglu, S., Bat, L., Kideys, A.E., Tugrul, S., Zagorodnyaya, J., Gündogdu, A., Akbulut, M., Çulha, M., Gönlügür, G., Eker, E., ve Satilmis, H.H., (2002). Orta Karadeniz’in Sinop Burnu Bölgesinin Biyokimyasal Dönüsüm Çalismalari, TÜBITAK Proje No: YDABÇAG- 619/G 197Y156. 92s.
- nCaddy, J. F., (1993). Toward a cooperative evaluation human impacts on fishery ecosystems and semi-enclosed seas. Fishery Science. 1(1): 57-95.
- nCaddy, J. and Griffiths, R., (1990), A Perspective on Recent Fishery Releted Events in the Black Sea, Studies and Review, General Fisheries Council for the Mediterranean, 63: 43-71.
- nCihangir, B. ve Uslu, B., (1992). Ege Denizi’nde Hamsi Baligi (Engraulis encrasicolus L., 1758))’nin Fekonditesi Üzerine Bir Ön Çalisma. Doga-Tr. J. Zoology, 16, 301-310.
- nCowan, V.G. and Houde, E.D., (1993). Relative Predation Potetials of Scypsomedusae, Ctenophores and Planktivorous Fish on Ichthiyoplankton in Chesapeake Bay. Mar. Ecol. 95: 55-65.
- nDaskalov, G.M., (2002). Overfishing Drives a Trophic Cascade in the Black Sea. Marine Ecology Progress Series 225: 53–63
- nDemir, N., (1959). Notes on the Variations of the Eggs of Anchovy (Engraulis encrasicholus CUV.) from Black, Marmara, Aegean and Mediterranean Seas. Publ. of Hidrobiol. Res. Inst., Faculty of Sciences, Univ. of Istanbul, Fen Fak. Hidrobiyoloji Arastirma Enstitüsü Yayinlari, Seri-B, Cilt IV, Sayi 4, Istanbul. 180-187.
- nDemir, N., (1965). Synopsis of Biological Data on Anchovy Engraulis encrasicolus (Linnaeus) 1758 (Mediterranean and adjacent seas). FAO Fisheries Synopsis No.26, Revission,1. SAST-Anchovy- 1,21(6),002,01. 42s.
- nEinarson, H. and Gürtürk, N., (1960). Abundance and Distribution of Eggs and Larvae of the Anchovy (Engraulis encrasicholus ponticus) In the Black Sea. Istanbul Üniv. Fen Fak. Mec. (B), 5 (1-2): 72-94.
- nEker E., L. Georgieva, L. Senichkina and A. E. Kideys, (1999). Phytoplankton distribution in the western and eastern Black Sea in spring and autumn 1995. ICES Journal of Marine Science 56: 15-22.
- nEker, E., (1999). Abundance and biomass of micro and nanophytoplankton in the northwestern and southern Black Sea in 1995. Master Thesis, Middle East Technical University, IMS, Erdemli, Icel, Turkey, 212 pp.
- nErgün, G., (1994). Distribution of five Calanoid copepod species in the southern Black Sea. M.S. Thesis, IMS-Middle East Technical University/Ankara, Turkey, 117pp.
- nErkan, F., Gücü, A. C. and Zagorodnyaya, J., (2000). The diel vertical distribution of zooloplankton in the southeast Black Sea. Turk. J. Zool. 24: 417-427.
- nErkoyuncu, I. ve Özdamar, E., (1989). Estimation of the Age, Size and Sex Composition and Growth Parameters of Anchovy Engraulis encrasicolus (L) in the Black Sea. Fisher. Res., 7: 241-247.
- nErüz, Ç., Durukanoglu, H.F. ve Bahar, M., (1993). Mezuniyetin Ilk Yilinda Dogu Anadolu Bölgesi I. Su Ürünleri Sempozyumu, 23-25
- nHaziran 1993, Atatürk Üniv., Ziraat Fak., Su Ürünleri Bölümü, s 651- 659.
- nEzurum. Feyzioglu, A. M., (1990). Dogu Akdeniz fitoplankton türlerinin kalitatif ve kantitatif yönden arastirilmasi (Yüksek Lisans Tezi), KTÜ, 52 s.
- nFeyzioglu, A. M., (1996). Dogu Karadeniz Kiyisal Ekosisteminde Fitoplankton Dinamigindeki Mevsimsel Degisimler. (Doktora Tezi), Karadeniz Teknik Üniversitesi Fen Bil. Enst. Trabzon.
- nFinenko, G.A., Anninsky, B.E., Romanova, Z.A., Abolmasova, G.I. and Kideys, A.E., (2001). Chemical composition, respiration and feeding rates of the new alien ctenophore, Beroe ovata, in the Black Sea. Hydrobiology, 451: 177-186.
- nFinenko, G.A., Romanova, Z.A., Abolmasova, G.I., Anninsky, B., Svetlichny, L.S., Hubareva, E.S., Bat, L. and Kideys, A.E., (2003). Population Dynamics, Ingestion, Growth, and Reproduction Rates of the Invader Beroe ovata and Its Impact on Plankton Community in Sevastopal Bay, the Black Sea, Journal of Plankton research, 25(5): 539-549.nnGerlach, S., (1990). Nitrogen, Phosphorous, Plankton and Oxygen deficiency in the German Bight and Kiel Bay. Kieler Meeresforchungen, Sonderheft Nr. No.7:341 p.nnGordina, A.D., Subbotin, A.A. and Klimova, T.N., (1990). Abundance and Peculiarities of Ichthyoplankton Distribution in the Western Black Sea in Summer Period 1988. Ukr. Acad. Sci. Kavalevsky Institute of Biologoy of the Southern Seas. Sevastopol. 32p.Dep. in VINITI 19.10.90, N 5410-B90 (In Russian)nnGordina, A.D., Zagorodnyaya, J.A., Kideys, A.E., Bat, L. and Satilmis, H.H., (2005). Summer Ichthyoplankton, Food Supply of Fish Larvae and Impact of Invasive Ctepnohores on the Nutrition of Fish Larvae in the Black Sea During 2000- 2001. J. Mar. Biol. Ass. U.K., 85: 537- 548.nnGözler, A. M. ve Çiloglu, E., (1998). Rize- Hopa Açiklarinda 1997-1998 Avlanma Sezonunda Avlanan Hamsi (Engraulis Encrasicolus L., 1758) Baligi’nin Bazi Populasyon Parametereleri Üzerine Bir Arastirma. III. Su Ürünleri Sempozyumu, 10-12 Haziran 1998,nnErzurum. Gubanova, A., (2000). Occurence of A. tonsa Dana in the Black Sea. Was it introduced from the Mediterranean? Mediterranean Marine Science, 1 (1): 105-109nnGubanova, A. D., Prusova, I. Yu, Nierman, U., Shadrin, N. V. and Polikarpov, I. G., (2001). Dramatic changes in the copepod community in Sevastopol Bay (Black Sea) during two decades (1976-1996). Senckenbergiana maritima, 31 (1): 17-27nnGucu, A.C., (1997). Role of fishing on Black Sea Ecosystem in Sensitivity to Change: Black Sea, Baltic Sea and North Sea, E. Özsoy and A. Mikaelyan (eds.), NATOASI Series, The Netherlands, Kluwer Academic Publishers, pp. 149-162nnGucu, A.C., (2002). Can Overfishing be Responsible for the Successful Establishment of Mnemiopsis leidyi in the Black Sea? Estuarine, Coastal and Shelf Science 54: 439-451pnnGürtürk, N., (1962). Plankton arastirmalarinin önemi. Balik ve Balikçilik Mecmuasi, X(11-12): 14-17.nnKaraçam, H. ve Düzgünes, E., (1990). Trabzon sahil seridi fitoplanktonu üzerine bir arastirma, I. Ü. Su Ürün. Dergisi, 4(1): 95-102nnKarnivola, V.P., (1955). Nabludenie za Rostom lichinok i malodi Azavskoi Khamsi v 1953 g Trud. Azov.-Chernomorsk.nauch.- issled. Inst.morsk.ryb.Khoz. (16) (In: nnDemir, N., 1965. Synopsis of Biological Data on Anchovy Engraulis encrasicolus (Linnaeus) 1758 (Mediterranean and adjacent seas). FAO Fisheries Synopsis No.26, Revission,1. SAST-Anchovy- 1,21(6),002,01. 42snnKideys, A. E., (1994). Recent dramatic changes in the Black Sea ecosystem: The reason for the sharp decrease in Turkish anchovy fisheries. Journal of Marine Systems 5: 171-181nnKideys, A. E., (2002). Fall and rise of the Black Sea ecosystem. Science’s Compass, vol 297: 1482-1483.nnKideys, A. E. and Romanova, Z., (2001). Distribution of gelatinous macrozooplankton in the southern Black Sea during 1996-1999. Marine Biology 139: 535-547nnKideys, A.E., Gordina, A. D., Niermann, U., Uysal, Z., Shiganova, T.A. and Bingel, F., (1998). Distribution of Eggs and Larvae of Anchovy with Respect to Ambient Conditions in the Southern Black Sea During 1993 and 1996. Ivanov, L.I. and Oguz, T. (eds) Ecosystem Modelling as a Management Tool for the Black Sea, 1: 189-198nnKideys, A. E., Gordina, A., Bingel, F. and Niermann, U., (1999). The effect of environmental conditions on the distribution of eggs and larvae of anchovy (Engraulis encrasicolus L.) in the Black Sea. ICES Journal of Marine Science 56: 58-64nnKideys, A. E., Kovalev, A. V., Shulman, G., Gordina, A. and Bingel, F., (2000). A review of zooplankton investigations of the Black Sea over the last decade. Journal of Marine Systems 24: 355-371.nnKonsulov, A. and Kamburska, L., (1998). Black Sea zooplankton structural dynamic and variability off the Bulgarian Black Sea coast during 1991-1995. In: NATO TUBlack Sea Project: Ecosystem Modeling as a Management Tool for the Black Sea, Symposium on Scientific Results, L. Ivanov & T. Oguz (eds.), Kluwer Academic Publishers, pp.281-292nnKoblentz-Mishke, 0. J., Volkovinsky, V V. and Kabanova, Yu. G. (1970). Plankton primary production in the World ocean. In: Wooster, W. (ed.) Scientific exploration of the South Pacific Ocean. Nat. Acad. Sci, Washington, D. C. p. 183-193nnKovalev A.V., Melnikov, V.V., Ostrovskaya N.A. and Prusova, I.Y., (1993). Zooplankton: Macroplankton, in, Plankton of the Black Sea.-Kiev: Naukova dumka,: pp. 183- 193 (in Russian)nnKovalev, A.V., Gubanova A.D., Kideys, E.A., Melnikov, V.V., Niermann, U., Ovstrovskaya, N.A., Prosova, I.Y., Skryabin V.A., Uysal, Z. and Zagoradnyaya J.A., (1998a). Long-term Changes in the Biomass and Composition of Fodder zooplankton in Coastal Regions of the Black Sea During the Period 1954 and 1996. In Ivanov, L.I. and Oguz, T. (eds) Ecosystem Modelling as a Management Tool for the Black Sea, 1: 209-219nnKovalev, A., Besiktepe, S., Zagorodnyaya, Yu. A. and Kideys, A. E., (1998b). Mediterranization of the Black Sea zooplankton is continuing. In: NATO TUBlack Sea Project: Ecosystem Modeling as a Management Tool for the Black Sea, Symposium on Scientific Results, L. Ivanov & T. Oguz (eds.), Kluwer Academic Publishers, pp.199-208.nnMihnea, P. E., (1992). Conventional methods applied in pollution control of the Romanian coastal waters of the Black Sea. In: Vollenweider, R. A., Marchetti, R., Viviani, R. (eds) Marine Coastal Eutrophication, Elsevier, New York, p. 1165 -1178nnMihnea, P.E. (1985). Effect of pollution on phytoplankton species. Rapp. Comm. Int. Mer. Medit., 29 (9): 85-88. Moncheva, S. P., (1991). Phytoplankton under the influence of eutrophicationpecularities and ecological significance. Karadeniz Egitim Kültür ve Çevre Koruma Vakfi Sempozyumu, Istanbul Atatürk Kültür Merkezi, 16-18 Eylül 1991, TurkeynnMurray, J. M., Codispoti, L. A. and Friederich, G. E., (1993). Redox environments: The suboxic zone in the Black Sea. In: Huang, C. P. O’Melia, R. O., Morgan, J. J. (eds) Aquatic Chemistry, American Chemical SocietynnMutlu, E., Bingel, F., Gucu, A.C., Melnikov, V.V., Niermann, U., Ostr, N.A. and Zaika, V.E., (1994). Distribution of the new invader Mnemiopsis sp. and the resident Au relia aurita and Pleurobrachia pileus populations in the Black Sea in the years 1991-1993. ICES Journal of Marine Science 51: 407-421nnNiermann, U., Bingel, F., Gorban, A., Gordina, A. D., Gücü, A. C., Kideys, A. E., Konsulov, A., Radu, G., Subbotin, A. A. and Zaika, V. E., (1994). Distribution of anchovy eggs and larvae (Engraulis encrasicolus Cuv.) in the Black Sea in 1991- 1992. ICES J. of Marine Science 51, 395- 406nnNiermann, U., Bingel, F., Ergün, G. and Greve, W. (1998). Fluctation of Dominant Mesozooplankton Species in the Black Sea, Nort Sea and Baltic Sea: Is General Trend Recognisable? Turkish J. Zoology, 22, 63-81nnNiermann, U., Kideys, A.E., Kovalev, A.V., Melnikov, V. and Belokopytov, V.V., (1999). Fluctuations of Pelagic Species of the Open Black Sea During 1980-1995 and Possible Teleconnections. In: S. Besiktepe et al. (Eds.). Environmental Degradation of the Black Sea: Challenges and Remedies. Kluwer Academic Publishers: 147–173nnNikolsky, G.V., 1957. (Hentschel, F.J. Transl.) Spezielle Fischkunde. Berlin, Deutscher Verlag der Wissenschaften. Oguz, T., (2005b). Black Sea Ecosystem Response to Climatic Teleconnections Oceonography, 18: 118-129nnOguz, T., P. La Violette and U. Unluata, (1992)."Upper layer circulation of the southern Black Sea: Its variability as inferred from hydrographic and satellite observations". J. Geophys. Research, 97(C8): 12569-12584nnOguz, T., Latun, V. S., Latif, M. A., Vladimirov, V. V., Sur, H. I., Markov, A. A., Özsoy, E., Kotovshchikov, V. V, Eremeev, V. V. and Ünlüata, Ü., (1993). Circulation in the surface and intermediate layers of the Black Sea. Deep-Sea Research I. 40: 1597-1612nnOguz, T., Ducklow, H., Malanotte-Rizzoli, P., Tugrul, S., Nezlin, N. P. and Ünlüata Ü., (1996). Simulation of annual plankton productivity cycle in the Black Sea by a one dimentional physical-biological model model. Journal of Geophysical Research. 101(C7): 16585-16599nnOguz, T., (2005a). Long-term Impacts of Antropogenic Forcing on the Black Sea Ecosystem, Oceonography, 18: 104-113nnÖzel, I., (1998). Planktonoloji (Cilt I). Plankton ekolojisi ve arastirma yöntemleri. Ege Üniversitesi Su Ürünleri Fak. Yayinlari, No:56, 271nnÖzsoy, E., Oguz, T., Latif, M. A. and Ünlüata, Ü., (1986). Oceanography of the Turkish Straits- first annual report, vol. I. Physical Oceanography of the Turkish Straits. Institute of Marine Science, METU, Içel, Turkey, p. 223nnÖzsoy, E., Ünlüata, Ü. and Z. Top, (1993). The Mediterranean Water Evolution, Material Transport by Double Diffusive Intrusions, and Interior Mixing in the Black Sea, Prog. Oceanogr., 31: 275-320nnÖzsoy, E. and S. Besiktepe, (1995). Sources of Double Diffusive Convection and Impacts on Mixing in the Black Sea, pp. 261-274, in: Brandt, A. and H. J. S. Fernando (editors), Double-Diffusive Convection, Geophysical Monograph 94, American Geophysical Union, 334 ppnnPavlovskaya, R. M., (1955). Vyzchivanie Chernomorski Khamsy na rannikh etapakh razvitiia. Trud.azov.-chernomorsk nach.- issled.Inst.morsk.ryb. Khoz., (16): 99-121 (In: Demir, N., 1965. Synopsis of Biological Data on Anchovy Engraulis encrasicolus (Linnaeus) 1758 (Mediterranean and adjacent seas). FAO Fisheries Synopsis No.26, Revission,1. SAST-Anchovy-1,21(6),002,01. 42snnPetrova- Karadjova, V., (1973). Dynamics of the biomass of phytoplankton in the Black Sea off the Bulgarian coast during the period of 1964-1970. Proc. Res. Inst. Ocean. Fish., 12: 41-66nnRoss, D. A., Uchupi, E., Prada, K. E. and Macilaine, J. C., (1974). Bathymetry and Microtopography of the Black Sea. In: Degens, T and Ross, A. (eds) The Black Sea geology, chemistry and biology. American Association of Petroleum Geologists, p. 1-10nnSamsun, O., Samsun, N. and Karamollaoglu, A.C., (2004). Age, Growth, and Mortality Rates of the European Anchovy (Engraulis encrasicolus L. 1758) off the Turkish Black Sea Coast. Turk J. Vet. Anim. Sci., 28: 901-910nnSatilmis, H.H., (2001). Balik Yumurta ve Larvalarinin Sinop Yarimadasi’nda Mevsimsel Olarak Dagilimi. Ondokuz Mayis Üniv. Su Ürünleri Yetistiriciligi Anabilim Dali, Yüksek Lisan Tezi. 82snnSatilmis, H.H., (2005). Sinop Kiyilarinda Küçük Pelajik Baliklarin Yumurta Üretimi ile Yumurta ve Larvalarinin Dagilimi. Ondokuz Mayis Üniversitesi Fen Bilimleri Enst. Su Ürünleri Avlama ve Isleme Teknolojisi Anabilim Dali, Doktora Tezi, Samsun, 160snnShiganova, T.A., (1998). Invasion on the Black Sea by the ctenophore Mnemiopsis leidyi and recent changes in pelagic community structure, Fisheries Oceanography, 7, 305- 310nnShiganova, T.A., (2004). Some results of studying the intruder Mnemiopsis leidyi (A. Agassiz) in the Black Sea Ctenophore Mnemiopsis leidyi (A. Agassiz) in the Azov and Black Sea: its biology and concequences of its intrusion, TUDAV publications, Istanbul, 28-68, 975-8825- 00-3nnShiganova, T.A. and Bulgakova, Y.V., (2000). Effects of gelatinous plankton on Black Sea and sea of Azov fish and their food resources, ICES Journal of Marine Science 57nnShiganova, T.A., Kideys, A.E., Gucu A.C., Niermann, U. and Khoroshilov, V.S., (1998). Changes in Species Diversity and Abundance of the Main Components of the Black Sea Pelagic Community During the Last Decade. In: NATO TU-Black Sea Project: Ecosystem Modeling as a Management Tool for the Black Sea, Symposium on Scientific Results, L. Ivanov & T. Oguz (eds.), Kluwer Academic Publishers, pp.171-188nnShiganova, T.A., Bulgakova, U.V., Sorokin, P.U. and Lukashev, U.F., (2000a). Results of study of the new invader Beroe ovata in the Black Sea, Izv ISR. Ser. Biology No 2 p 248-256nnShiganova, T.A., Bulgakova, U.V., Volovik, S.P., Mirzoyan, Z.A. and Dudkin, S.I., (2000b). New invader Beroe ovata and Its impact on the Ecosystem of Azov-Black Sea basin in August-September 1999// Ctenophore Mnemiopsis leidyi (A.Agassiz) in Azov and Black Seas and Consequences of its Introduction. Editd by S.P.Volovik. Rostov-na-Donu. p 432-449nnShiganova, T.A.,Kamakin, A.M., Zhukova, O.P., Ushivtsev, V.B., Dulimov, A.B. & Musaeva, E.I., (2001). The invader into the Caspian Sea Ctenophore Mnemiopsis leidyi and Its Initial Effect on the Pelajik Ecosystem, Oceanology, 41: 517-524nnShiganova, T.A., Musayeva, E.I., Bulgakova, Y.V., Mirzoyan, Z.A. and Martynyuk, M.L., (2003). Invaders ctenophores Mnemiopsis leidyi (A. Agassiz) and Beroe ovata Mayer 1912, and their influence on the pelagic ecosystem of Northeastern Black Sea. Biology Bulletin, 30: 180-190nnShiganova, T.A., Bulgakova, Y.V., Dumond, H., Mikaelyan, A., Glazov, D.M., Bulgakova, Y.V., Musayeva E.I., Sorokin P.Y., Pautova, L.A., Mirzoyan, Z.A. and Studenikina, E.I., (2004a). Interactions Between the Invading Ctenophores Mnemiopsis leidyi (A. Agasiz) and Beroe ovata Mayer 1912 and Influence on Pelajik Ecosystem of the Northeastern Black Sea, Aquatic Invasion in the Black, Caspian and Mediterranean Seas, 33-70nnShiganova, T.A., Bulgakova, Y.V., Dumond, J.H., Mikaelyan, A., Glazov, D.M., Bulgakova, Y.V., Musayeva, E.I., Sorokin, P.Y., Pautova, L.A., Mirzoyan, Z.A. and Studenikina, E.I., (2004b). Population dynamics of Mnemiopsis leidyi in the Caspian Sea, and effects on the Caspian Sea, Aquatic Invasion in the Black, Caspian and Mediterranean Seas, pp.71- 107nnShulman, G. and Yuneva, T.V., (2002). Fishes as indicator of state of the Black Sea and Mediterranean ecosystems. In:Yilmaz A.E; Salihoglu, I; Mutlu, E.;(eds). Oceanograpyof the eastern Mediterranean and Black Sea. Abstracts, 2nd International Conferance METU, Ankara, p:175nnSlastenenko, E., (1955-1956). Karadeniz Havzasi Baliklari, Et ve Balik Kurumu Umum Müdürlügü Yayinlarindan, Istanbul, 711s. Sorokin, Yu. I., (1983). The Black Sea. In: Ketchum, B. H. (eds) Ecosystems of the world estuaries and enclosed seas. Elsevier, Amsterdam, p. 253-291nnSukhanova, I. N., Georgieva, L. V., Mikaelyan, A. S., Sergeeva, O. V., (1988). Phytoplankton of the open waters of the Black Sea in late springi, in Sovremennoye sostoyaniye ekosistemy Chernogo morya (Modern State of the Black Sea Ecosystem), Nauka, Moskow, p. 86-97nnSullivan, B. K., and L. T. Mcmanijs. (1986). Factors controlling seasonal succession of copepods Acartia hudsonica and A. tonsa in Narragansett Bay, Rhode Island: Temperature and resting egg production. Mar. Ecol. Prog. Ser. 28: 121-128nnSur, I.H., Ozsoy, E. and Unluata, U., (1993). Boundary Current Instabilities, Upwelling., Shelf Mixing and Eutrophication Processes in the Black Sea, Institute of Marine Sciences, ODTÜ, ErdemlinnSur, H. I., Özsoy, E. and Ünlüata, Ü., (1994). Boundary current instabilities, upwelling, shelf mixing and eutrophication processes in the Black sea. Prog. Oceanog., 33: 249- 302nnTkach, A. V., Gordina, A. D., Kideys, A. E., Niermann, U. and Zaika, V. E., (1998). Changes in the larval nutrition of the Black Sea fishes with respect to plankton. In: NATO TU-Black Sea, Project: Ecosystem Modeling as a Managment Tool for the Black Sea, Symposium on Scientific Results, L. Ivanov & T. Oguz (eds.), Kluwer Academic Publishers, 1: 235-248nnTolmazin, D., (1985). Changing coastal oceanography of the Black Sea. I: Northwestern Shelf., Prog. Oceanog., 15: 217-276nnTsikhon-Lukanina, E.A., Reznichenko, O.G., Lukasheva, T.A., (1991), Quantitative patterns of feeding of the Black Sea ctenophore Mnemiopsis leidyi, Oceanology, 31:196-199nnTsikhon-Lukanina, E.A., Reznichenko, O.G. and Lukasheva, T.A., (1993). Level of fish fry consumption by Mnemiopsis in the Black Sea shelf. Okeanologiya 33: 895- 899nnTürkoglu, M. and T. Koray., (2002). Species Succession and Diversity of Phytoplankton in the Neritic Waters of Southern Black Sea (The Bay of Sinop, Turkey). Turk. J. Bot. 26: 235-252nnTürkoglu, M., (1998). Orta Karadeniz Bölgesi’nin Sinop yarimadasi kiyilari fitoplankton kompozisyonu ve degisimini etkileyen faktörler. E. Ü. Fen Bil. Enst. (Doktora Tezi)nnIzmir. Ünsal, N., (1989). Karadeniz’deki Hamsi Baligi, Engraulis encrasicolus (L, 1758)’nin Yas- Boy- Agirlik Iliskisi ve En Küçük Av Büyüklügünün Saptanamsi üzerine Bir Arastirma. IÜ Su Ürünleri Dergisi, 3 (1-2): 17-18nnUysal, Z., (1993). A preliminary study on some plankters along The Turkish Black Sea Coast- Species composition and spatial distribution. ODTÜ, Den. Bil. Enst. (Doktora Tezi), 138pnnUysal, Z., Kideys. A. E., L. Senichkina., L. Georgieva., D. Altukhov., L. Kuzmenko., L. Manjos., E. Mutlu., and E. Eker., (1997). Phytoplankton patches formed along the southern Black Sea coast in the spring and summer of 1996. NATO TUBlack Sea Assessment Workshop. 15-19 June 1997, Crimea, UkrainennUysal, Z. and Sur, H. I., (1995). Net phytoplankton discriminating patches along the Southern Black Sea coast in winter 1990. Oceanologica Acta, 18/6: 639-647nnÜnal, E., (2002). Seasonality of zooplankton in the Southern Black Sea in 1999 and Genetics of Calanus euxinus (Copepoda). M.S. Thesis, IMS-Middle East Technical University/Ankara, Turkey, 214ppnnÜnlüata, U., T. Oguz, M.A. Latif and E. Ozsoy., (1990). "On the physical oceanography of the Turksh Straits." In The Physical Oceanography of the Sea Straits, L.J. Pratt (Ed.). NATO/ASI Series, 318: 25-60, nnKluwer Academic Publishers, Dordrecht. Üstün, F., (2005). Karadeniz’in Sinop Burnu Bölgesinin Zooplankton Kompozisyonu ve Mevsimlse Dagilimi, Fen Bilimsleri Enstitüsü Su Ürünleri Temel Bilimleri Anabilim Dali, Yüksek Lisans Tezi, 173 snnSamsun. Vinagradov M.E, , Shushkina E.A, Musayeva E.I, Sorokin P.Yu, (1989). A new acclimated species in the Black Sea: the ctenophore Mnemiopsis leidyi (Ctenophore: Lobata). Oceanology 29: 220-224.nnVinogradov, M.E. and Flint, M.V., (1985). Researches of the Pelagical Ecosystem of the Black Sea: 6th Cruise of R/V “Vityaz” (April, 19-June, 3 1984). Oceanology, 25: 168-171nnVinogradov, M.E. and Shushkina E.A., (1992). Temporal change in community structure in the open Black Sea. Oceanology, 32: 485-491nnVinogradov, M.E., (1987). Modern Tendencies of Changing of the Black Sea Ecosystem, Vestnik AN SSSR 7 (10): 56-57 (in Russian)nnVladimirov, V.L., Mankovsky,V.I., Solov’ev, M.V., Mishonov. A.V. and Besiktepe, S.T., (1999). Hydro-Optical studies of the Black sea: History and status, Environmental Degration Of the Black Sea: Challenges and Remeddies, 245-256nnVodyanitskii, V.A. and Kazanova, I., (1954). Opredelitel Pelagicheskii Lichinok ryb Chernogomoria (Key to do Pelagic Fish Eggs and Larvae of the Black Sea). Tr. Vses.naucho-Issled. Inst. Morsk. Rybn. Khoz. Okeanogr., 28: 240-325 (in Russian)nnYildiz, N., (1997). Sinop ili iç liman pelajik Copepoda (Crustacea) faunasi üzerine bir arastirma. O.M.Ü. Fen Bilimleri Ens. (Yüksek Lisans Tezi), SamsunnnYüksek, A., (1993). Marmara Denizi’nin Kuzey Bölgesinde Teleostat Baliklarin Pelajik Yumurta ve Larvalarinin Dagilimi ve Bollugu (Bakirköy Marmara Ereglisi), Doktora Tezi, I.Ü. Deniz Bilimleri ve Isletmeciligi Ens., Istanbul, 1993nnYüksek, A. ve Gücü, A.C., (1994). Balik Yumurtalari Tayini Için Bir Bilgisayar Yazilimi (Karadeniz Pelajik Yumurtalari), Karadeniz Egitim-Kültür ve Çevre Koruma Vakfi, Istanbul, 51s. Zaitsev, YU. P. (1992). Recent changes in the trophic structure of the Black Sea. Fisheries Oceanography 1: 180-189nnZaitsev, Yu. P. and Aleksandrov, B. G., (1997). Recent man-made changes in the Black Sea ecosystem. In: Sensitivity to Change: Black Sea, Baltic Sea and North Sea, E. Ozsoy and A. Mikaelyan, editors, Kluwer Acad. Publ., pp. 25-32nnZaitsev, Yu. P. and Alexandrov, B. G., (1995). Recent man-made changes in the Black Sea ecosystem. Sensitivity of North Sea. Baltic Sea and Black Sea to antropogenic and climatic changes. Workshop, Varna- Bulgaria, 14-18 NovnnZhorov, V.A. and Boguslavsky, S.G., (1985). Tendencies of Some Hydrological- Hydrochemical Processes of the Black Sea, Meteorology and Hydrology 11: 63- 69.n
- nGerlach, S., (1990). Nitrogen, Phosphorous, Plankton and Oxygen deficiency in the German Bight and Kiel Bay. Kieler Meeresforchungen, Sonderheft Nr. No.7:341 p.
- nGordina, A.D., Subbotin, A.A. and Klimova, T.N., (1990). Abundance and Peculiarities of Ichthyoplankton Distribution in the Western Black Sea in Summer Period 1988. Ukr. Acad. Sci. Kavalevsky Institute of Biologoy of the Southern Seas. Sevastopol. 32p.Dep. in VINITI 19.10.90, N 5410-B90 (In Russian)
- nGordina, A.D., Zagorodnyaya, J.A., Kideys, A.E., Bat, L. and Satilmis, H.H., (2005). Summer Ichthyoplankton, Food Supply of Fish Larvae and Impact of Invasive Ctepnohores on the Nutrition of Fish Larvae in the Black Sea During 2000- 2001. J. Mar. Biol. Ass. U.K., 85: 537- 548.
- nGözler, A. M. ve Çiloglu, E., (1998). Rize- Hopa Açiklarinda 1997-1998 Avlanma Sezonunda Avlanan Hamsi (Engraulis Encrasicolus L., 1758) Baligi’nin Bazi Populasyon Parametereleri Üzerine Bir Arastirma. III. Su Ürünleri Sempozyumu, 10-12 Haziran 1998,
- nErzurum. Gubanova, A., (2000). Occurence of A. tonsa Dana in the Black Sea. Was it introduced from the Mediterranean? Mediterranean Marine Science, 1 (1): 105-109
- nGubanova, A. D., Prusova, I. Yu, Nierman, U., Shadrin, N. V. and Polikarpov, I. G., (2001). Dramatic changes in the copepod community in Sevastopol Bay (Black Sea) during two decades (1976-1996). Senckenbergiana maritima, 31 (1): 17-27
- nGucu, A.C., (1997). Role of fishing on Black Sea Ecosystem in Sensitivity to Change: Black Sea, Baltic Sea and North Sea, E. Özsoy and A. Mikaelyan (eds.), NATOASI Series, The Netherlands, Kluwer Academic Publishers, pp. 149-162
- nGucu, A.C., (2002). Can Overfishing be Responsible for the Successful Establishment of Mnemiopsis leidyi in the Black Sea? Estuarine, Coastal and Shelf Science 54: 439-451p
- nGürtürk, N., (1962). Plankton arastirmalarinin önemi. Balik ve Balikçilik Mecmuasi, X(11-12): 14-17.
- nKaraçam, H. ve Düzgünes, E., (1990). Trabzon sahil seridi fitoplanktonu üzerine bir arastirma, I. Ü. Su Ürün. Dergisi, 4(1): 95-102
- nKarnivola, V.P., (1955). Nabludenie za Rostom lichinok i malodi Azavskoi Khamsi v 1953 g Trud. Azov.-Chernomorsk.nauch.- issled. Inst.morsk.ryb.Khoz. (16) (In:
- nDemir, N., 1965. Synopsis of Biological Data on Anchovy Engraulis encrasicolus (Linnaeus) 1758 (Mediterranean and adjacent seas). FAO Fisheries Synopsis No.26, Revission,1. SAST-Anchovy- 1,21(6),002,01. 42s
- nKideys, A. E., (1994). Recent dramatic changes in the Black Sea ecosystem: The reason for the sharp decrease in Turkish anchovy fisheries. Journal of Marine Systems 5: 171-181
- nKideys, A. E., (2002). Fall and rise of the Black Sea ecosystem. Science’s Compass, vol 297: 1482-1483.
- nKideys, A. E. and Romanova, Z., (2001). Distribution of gelatinous macrozooplankton in the southern Black Sea during 1996-1999. Marine Biology 139: 535-547
- nKideys, A.E., Gordina, A. D., Niermann, U., Uysal, Z., Shiganova, T.A. and Bingel, F., (1998). Distribution of Eggs and Larvae of Anchovy with Respect to Ambient Conditions in the Southern Black Sea During 1993 and 1996. Ivanov, L.I. and Oguz, T. (eds) Ecosystem Modelling as a Management Tool for the Black Sea, 1: 189-198
- nKideys, A. E., Gordina, A., Bingel, F. and Niermann, U., (1999). The effect of environmental conditions on the distribution of eggs and larvae of anchovy (Engraulis encrasicolus L.) in the Black Sea. ICES Journal of Marine Science 56: 58-64
- nKideys, A. E., Kovalev, A. V., Shulman, G., Gordina, A. and Bingel, F., (2000). A review of zooplankton investigations of the Black Sea over the last decade. Journal of Marine Systems 24: 355-371.
- nKonsulov, A. and Kamburska, L., (1998). Black Sea zooplankton structural dynamic and variability off the Bulgarian Black Sea coast during 1991-1995. In: NATO TUBlack Sea Project: Ecosystem Modeling as a Management Tool for the Black Sea, Symposium on Scientific Results, L. Ivanov & T. Oguz (eds.), Kluwer Academic Publishers, pp.281-292
- nKoblentz-Mishke, 0. J., Volkovinsky, V V. and Kabanova, Yu. G. (1970). Plankton primary production in the World ocean. In: Wooster, W. (ed.) Scientific exploration of the South Pacific Ocean. Nat. Acad. Sci, Washington, D. C. p. 183-193
- nKovalev A.V., Melnikov, V.V., Ostrovskaya N.A. and Prusova, I.Y., (1993). Zooplankton: Macroplankton, in, Plankton of the Black Sea.-Kiev: Naukova dumka,: pp. 183- 193 (in Russian)
- nKovalev, A.V., Gubanova A.D., Kideys, E.A., Melnikov, V.V., Niermann, U., Ovstrovskaya, N.A., Prosova, I.Y., Skryabin V.A., Uysal, Z. and Zagoradnyaya J.A., (1998a). Long-term Changes in the Biomass and Composition of Fodder zooplankton in Coastal Regions of the Black Sea During the Period 1954 and 1996. In Ivanov, L.I. and Oguz, T. (eds) Ecosystem Modelling as a Management Tool for the Black Sea, 1: 209-219
- nKovalev, A., Besiktepe, S., Zagorodnyaya, Yu. A. and Kideys, A. E., (1998b). Mediterranization of the Black Sea zooplankton is continuing. In: NATO TUBlack Sea Project: Ecosystem Modeling as a Management Tool for the Black Sea, Symposium on Scientific Results, L. Ivanov & T. Oguz (eds.), Kluwer Academic Publishers, pp.199-208.
- nMihnea, P. E., (1992). Conventional methods applied in pollution control of the Romanian coastal waters of the Black Sea. In: Vollenweider, R. A., Marchetti, R., Viviani, R. (eds) Marine Coastal Eutrophication, Elsevier, New York, p. 1165 -1178
- nMihnea, P.E. (1985). Effect of pollution on phytoplankton species. Rapp. Comm. Int. Mer. Medit., 29 (9): 85-88. Moncheva, S. P., (1991). Phytoplankton under the influence of eutrophicationpecularities and ecological significance. Karadeniz Egitim Kültür ve Çevre Koruma Vakfi Sempozyumu, Istanbul Atatürk Kültür Merkezi, 16-18 Eylül 1991, Turkey
- nMurray, J. M., Codispoti, L. A. and Friederich, G. E., (1993). Redox environments: The suboxic zone in the Black Sea. In: Huang, C. P. O’Melia, R. O., Morgan, J. J. (eds) Aquatic Chemistry, American Chemical Society
- nMutlu, E., Bingel, F., Gucu, A.C., Melnikov, V.V., Niermann, U., Ostr, N.A. and Zaika, V.E., (1994). Distribution of the new invader Mnemiopsis sp. and the resident Au relia aurita and Pleurobrachia pileus populations in the Black Sea in the years 1991-1993. ICES Journal of Marine Science 51: 407-421
- nNiermann, U., Bingel, F., Gorban, A., Gordina, A. D., Gücü, A. C., Kideys, A. E., Konsulov, A., Radu, G., Subbotin, A. A. and Zaika, V. E., (1994). Distribution of anchovy eggs and larvae (Engraulis encrasicolus Cuv.) in the Black Sea in 1991- 1992. ICES J. of Marine Science 51, 395- 406
- nNiermann, U., Bingel, F., Ergün, G. and Greve, W. (1998). Fluctation of Dominant Mesozooplankton Species in the Black Sea, Nort Sea and Baltic Sea: Is General Trend Recognisable? Turkish J. Zoology, 22, 63-81
- nNiermann, U., Kideys, A.E., Kovalev, A.V., Melnikov, V. and Belokopytov, V.V., (1999). Fluctuations of Pelagic Species of the Open Black Sea During 1980-1995 and Possible Teleconnections. In: S. Besiktepe et al. (Eds.). Environmental Degradation of the Black Sea: Challenges and Remedies. Kluwer Academic Publishers: 147–173
- nNikolsky, G.V., 1957. (Hentschel, F.J. Transl.) Spezielle Fischkunde. Berlin, Deutscher Verlag der Wissenschaften. Oguz, T., (2005b). Black Sea Ecosystem Response to Climatic Teleconnections Oceonography, 18: 118-129
- nOguz, T., P. La Violette and U. Unluata, (1992)."Upper layer circulation of the southern Black Sea: Its variability as inferred from hydrographic and satellite observations". J. Geophys. Research, 97(C8): 12569-12584
- nOguz, T., Latun, V. S., Latif, M. A., Vladimirov, V. V., Sur, H. I., Markov, A. A., Özsoy, E., Kotovshchikov, V. V, Eremeev, V. V. and Ünlüata, Ü., (1993). Circulation in the surface and intermediate layers of the Black Sea. Deep-Sea Research I. 40: 1597-1612
- nOguz, T., Ducklow, H., Malanotte-Rizzoli, P., Tugrul, S., Nezlin, N. P. and Ünlüata Ü., (1996). Simulation of annual plankton productivity cycle in the Black Sea by a one dimentional physical-biological model model. Journal of Geophysical Research. 101(C7): 16585-16599
- nOguz, T., (2005a). Long-term Impacts of Antropogenic Forcing on the Black Sea Ecosystem, Oceonography, 18: 104-113
- nÖzel, I., (1998). Planktonoloji (Cilt I). Plankton ekolojisi ve arastirma yöntemleri. Ege Üniversitesi Su Ürünleri Fak. Yayinlari, No:56, 271
- nÖzsoy, E., Oguz, T., Latif, M. A. and Ünlüata, Ü., (1986). Oceanography of the Turkish Straits- first annual report, vol. I. Physical Oceanography of the Turkish Straits. Institute of Marine Science, METU, Içel, Turkey, p. 223
- nÖzsoy, E., Ünlüata, Ü. and Z. Top, (1993). The Mediterranean Water Evolution, Material Transport by Double Diffusive Intrusions, and Interior Mixing in the Black Sea, Prog. Oceanogr., 31: 275-320
- nÖzsoy, E. and S. Besiktepe, (1995). Sources of Double Diffusive Convection and Impacts on Mixing in the Black Sea, pp. 261-274, in: Brandt, A. and H. J. S. Fernando (editors), Double-Diffusive Convection, Geophysical Monograph 94, American Geophysical Union, 334 pp
- nPavlovskaya, R. M., (1955). Vyzchivanie Chernomorski Khamsy na rannikh etapakh razvitiia. Trud.azov.-chernomorsk nach.- issled.Inst.morsk.ryb. Khoz., (16): 99-121 (In: Demir, N., 1965. Synopsis of Biological Data on Anchovy Engraulis encrasicolus (Linnaeus) 1758 (Mediterranean and adjacent seas). FAO Fisheries Synopsis No.26, Revission,1. SAST-Anchovy-1,21(6),002,01. 42s
- nPetrova- Karadjova, V., (1973). Dynamics of the biomass of phytoplankton in the Black Sea off the Bulgarian coast during the period of 1964-1970. Proc. Res. Inst. Ocean. Fish., 12: 41-66
- nRoss, D. A., Uchupi, E., Prada, K. E. and Macilaine, J. C., (1974). Bathymetry and Microtopography of the Black Sea. In: Degens, T and Ross, A. (eds) The Black Sea geology, chemistry and biology. American Association of Petroleum Geologists, p. 1-10
- nSamsun, O., Samsun, N. and Karamollaoglu, A.C., (2004). Age, Growth, and Mortality Rates of the European Anchovy (Engraulis encrasicolus L. 1758) off the Turkish Black Sea Coast. Turk J. Vet. Anim. Sci., 28: 901-910
- nSatilmis, H.H., (2001). Balik Yumurta ve Larvalarinin Sinop Yarimadasi’nda Mevsimsel Olarak Dagilimi. Ondokuz Mayis Üniv. Su Ürünleri Yetistiriciligi Anabilim Dali, Yüksek Lisan Tezi. 82s
- nSatilmis, H.H., (2005). Sinop Kiyilarinda Küçük Pelajik Baliklarin Yumurta Üretimi ile Yumurta ve Larvalarinin Dagilimi. Ondokuz Mayis Üniversitesi Fen Bilimleri Enst. Su Ürünleri Avlama ve Isleme Teknolojisi Anabilim Dali, Doktora Tezi, Samsun, 160s
- nShiganova, T.A., (1998). Invasion on the Black Sea by the ctenophore Mnemiopsis leidyi and recent changes in pelagic community structure, Fisheries Oceanography, 7, 305- 310
- nShiganova, T.A., (2004). Some results of studying the intruder Mnemiopsis leidyi (A. Agassiz) in the Black Sea Ctenophore Mnemiopsis leidyi (A. Agassiz) in the Azov and Black Sea: its biology and concequences of its intrusion, TUDAV publications, Istanbul, 28-68, 975-8825- 00-3
- nShiganova, T.A. and Bulgakova, Y.V., (2000). Effects of gelatinous plankton on Black Sea and sea of Azov fish and their food resources, ICES Journal of Marine Science 57
- nShiganova, T.A., Kideys, A.E., Gucu A.C., Niermann, U. and Khoroshilov, V.S., (1998). Changes in Species Diversity and Abundance of the Main Components of the Black Sea Pelagic Community During the Last Decade. In: NATO TU-Black Sea Project: Ecosystem Modeling as a Management Tool for the Black Sea, Symposium on Scientific Results, L. Ivanov & T. Oguz (eds.), Kluwer Academic Publishers, pp.171-188
- nShiganova, T.A., Bulgakova, U.V., Sorokin, P.U. and Lukashev, U.F., (2000a). Results of study of the new invader Beroe ovata in the Black Sea, Izv ISR. Ser. Biology No 2 p 248-256
- nShiganova, T.A., Bulgakova, U.V., Volovik, S.P., Mirzoyan, Z.A. and Dudkin, S.I., (2000b). New invader Beroe ovata and Its impact on the Ecosystem of Azov-Black Sea basin in August-September 1999// Ctenophore Mnemiopsis leidyi (A.Agassiz) in Azov and Black Seas and Consequences of its Introduction. Editd by S.P.Volovik. Rostov-na-Donu. p 432-449
- nShiganova, T.A.,Kamakin, A.M., Zhukova, O.P., Ushivtsev, V.B., Dulimov, A.B. & Musaeva, E.I., (2001). The invader into the Caspian Sea Ctenophore Mnemiopsis leidyi and Its Initial Effect on the Pelajik Ecosystem, Oceanology, 41: 517-524
- nShiganova, T.A., Musayeva, E.I., Bulgakova, Y.V., Mirzoyan, Z.A. and Martynyuk, M.L., (2003). Invaders ctenophores Mnemiopsis leidyi (A. Agassiz) and Beroe ovata Mayer 1912, and their influence on the pelagic ecosystem of Northeastern Black Sea. Biology Bulletin, 30: 180-190
- nShiganova, T.A., Bulgakova, Y.V., Dumond, H., Mikaelyan, A., Glazov, D.M., Bulgakova, Y.V., Musayeva E.I., Sorokin P.Y., Pautova, L.A., Mirzoyan, Z.A. and Studenikina, E.I., (2004a). Interactions Between the Invading Ctenophores Mnemiopsis leidyi (A. Agasiz) and Beroe ovata Mayer 1912 and Influence on Pelajik Ecosystem of the Northeastern Black Sea, Aquatic Invasion in the Black, Caspian and Mediterranean Seas, 33-70
- nShiganova, T.A., Bulgakova, Y.V., Dumond, J.H., Mikaelyan, A., Glazov, D.M., Bulgakova, Y.V., Musayeva, E.I., Sorokin, P.Y., Pautova, L.A., Mirzoyan, Z.A. and Studenikina, E.I., (2004b). Population dynamics of Mnemiopsis leidyi in the Caspian Sea, and effects on the Caspian Sea, Aquatic Invasion in the Black, Caspian and Mediterranean Seas, pp.71- 107
- nShulman, G. and Yuneva, T.V., (2002). Fishes as indicator of state of the Black Sea and Mediterranean ecosystems. In:Yilmaz A.E; Salihoglu, I; Mutlu, E.;(eds). Oceanograpyof the eastern Mediterranean and Black Sea. Abstracts, 2nd International Conferance METU, Ankara, p:175
- nSlastenenko, E., (1955-1956). Karadeniz Havzasi Baliklari, Et ve Balik Kurumu Umum Müdürlügü Yayinlarindan, Istanbul, 711s. Sorokin, Yu. I., (1983). The Black Sea. In: Ketchum, B. H. (eds) Ecosystems of the world estuaries and enclosed seas. Elsevier, Amsterdam, p. 253-291
- nSukhanova, I. N., Georgieva, L. V., Mikaelyan, A. S., Sergeeva, O. V., (1988). Phytoplankton of the open waters of the Black Sea in late springi, in Sovremennoye sostoyaniye ekosistemy Chernogo morya (Modern State of the Black Sea Ecosystem), Nauka, Moskow, p. 86-97
- nSullivan, B. K., and L. T. Mcmanijs. (1986). Factors controlling seasonal succession of copepods Acartia hudsonica and A. tonsa in Narragansett Bay, Rhode Island: Temperature and resting egg production. Mar. Ecol. Prog. Ser. 28: 121-128
- nSur, I.H., Ozsoy, E. and Unluata, U., (1993). Boundary Current Instabilities, Upwelling., Shelf Mixing and Eutrophication Processes in the Black Sea, Institute of Marine Sciences, ODTÜ, Erdemli
- nSur, H. I., Özsoy, E. and Ünlüata, Ü., (1994). Boundary current instabilities, upwelling, shelf mixing and eutrophication processes in the Black sea. Prog. Oceanog., 33: 249- 302
- nTkach, A. V., Gordina, A. D., Kideys, A. E., Niermann, U. and Zaika, V. E., (1998). Changes in the larval nutrition of the Black Sea fishes with respect to plankton. In: NATO TU-Black Sea, Project: Ecosystem Modeling as a Managment Tool for the Black Sea, Symposium on Scientific Results, L. Ivanov & T. Oguz (eds.), Kluwer Academic Publishers, 1: 235-248
- nTolmazin, D., (1985). Changing coastal oceanography of the Black Sea. I: Northwestern Shelf., Prog. Oceanog., 15: 217-276
- nTsikhon-Lukanina, E.A., Reznichenko, O.G., Lukasheva, T.A., (1991), Quantitative patterns of feeding of the Black Sea ctenophore Mnemiopsis leidyi, Oceanology, 31:196-199
- nTsikhon-Lukanina, E.A., Reznichenko, O.G. and Lukasheva, T.A., (1993). Level of fish fry consumption by Mnemiopsis in the Black Sea shelf. Okeanologiya 33: 895- 899
- nTürkoglu, M. and T. Koray., (2002). Species Succession and Diversity of Phytoplankton in the Neritic Waters of Southern Black Sea (The Bay of Sinop, Turkey). Turk. J. Bot. 26: 235-252
- nTürkoglu, M., (1998). Orta Karadeniz Bölgesi’nin Sinop yarimadasi kiyilari fitoplankton kompozisyonu ve degisimini etkileyen faktörler. E. Ü. Fen Bil. Enst. (Doktora Tezi)
- nIzmir. Ünsal, N., (1989). Karadeniz’deki Hamsi Baligi, Engraulis encrasicolus (L, 1758)’nin Yas- Boy- Agirlik Iliskisi ve En Küçük Av Büyüklügünün Saptanamsi üzerine Bir Arastirma. IÜ Su Ürünleri Dergisi, 3 (1-2): 17-18
- nUysal, Z., (1993). A preliminary study on some plankters along The Turkish Black Sea Coast- Species composition and spatial distribution. ODTÜ, Den. Bil. Enst. (Doktora Tezi), 138p
- nUysal, Z., Kideys. A. E., L. Senichkina., L. Georgieva., D. Altukhov., L. Kuzmenko., L. Manjos., E. Mutlu., and E. Eker., (1997). Phytoplankton patches formed along the southern Black Sea coast in the spring and summer of 1996. NATO TUBlack Sea Assessment Workshop. 15-19 June 1997, Crimea, Ukraine
- nUysal, Z. and Sur, H. I., (1995). Net phytoplankton discriminating patches along the Southern Black Sea coast in winter 1990. Oceanologica Acta, 18/6: 639-647
- nÜnal, E., (2002). Seasonality of zooplankton in the Southern Black Sea in 1999 and Genetics of Calanus euxinus (Copepoda). M.S. Thesis, IMS-Middle East Technical University/Ankara, Turkey, 214pp
- nÜnlüata, U., T. Oguz, M.A. Latif and E. Ozsoy., (1990). "On the physical oceanography of the Turksh Straits." In The Physical Oceanography of the Sea Straits, L.J. Pratt (Ed.). NATO/ASI Series, 318: 25-60,
- nKluwer Academic Publishers, Dordrecht. Üstün, F., (2005). Karadeniz’in Sinop Burnu Bölgesinin Zooplankton Kompozisyonu ve Mevsimlse Dagilimi, Fen Bilimsleri Enstitüsü Su Ürünleri Temel Bilimleri Anabilim Dali, Yüksek Lisans Tezi, 173 s
- nSamsun. Vinagradov M.E, , Shushkina E.A, Musayeva E.I, Sorokin P.Yu, (1989). A new acclimated species in the Black Sea: the ctenophore Mnemiopsis leidyi (Ctenophore: Lobata). Oceanology 29: 220-224.
- nVinogradov, M.E. and Flint, M.V., (1985). Researches of the Pelagical Ecosystem of the Black Sea: 6th Cruise of R/V “Vityaz” (April, 19-June, 3 1984). Oceanology, 25: 168-171
- nVinogradov, M.E. and Shushkina E.A., (1992). Temporal change in community structure in the open Black Sea. Oceanology, 32: 485-491
- nVinogradov, M.E., (1987). Modern Tendencies of Changing of the Black Sea Ecosystem, Vestnik AN SSSR 7 (10): 56-57 (in Russian)
- nVladimirov, V.L., Mankovsky,V.I., Solov’ev, M.V., Mishonov. A.V. and Besiktepe, S.T., (1999). Hydro-Optical studies of the Black sea: History and status, Environmental Degration Of the Black Sea: Challenges and Remeddies, 245-256
- nVodyanitskii, V.A. and Kazanova, I., (1954). Opredelitel Pelagicheskii Lichinok ryb Chernogomoria (Key to do Pelagic Fish Eggs and Larvae of the Black Sea). Tr. Vses.naucho-Issled. Inst. Morsk. Rybn. Khoz. Okeanogr., 28: 240-325 (in Russian)
- nYildiz, N., (1997). Sinop ili iç liman pelajik Copepoda (Crustacea) faunasi üzerine bir arastirma. O.M.Ü. Fen Bilimleri Ens. (Yüksek Lisans Tezi), Samsun
- nYüksek, A., (1993). Marmara Denizi’nin Kuzey Bölgesinde Teleostat Baliklarin Pelajik Yumurta ve Larvalarinin Dagilimi ve Bollugu (Bakirköy Marmara Ereglisi), Doktora Tezi, I.Ü. Deniz Bilimleri ve Isletmeciligi Ens., Istanbul, 1993
- nYüksek, A. ve Gücü, A.C., (1994). Balik Yumurtalari Tayini Için Bir Bilgisayar Yazilimi (Karadeniz Pelajik Yumurtalari), Karadeniz Egitim-Kültür ve Çevre Koruma Vakfi, Istanbul, 51s. Zaitsev, YU. P. (1992). Recent changes in the trophic structure of the Black Sea. Fisheries Oceanography 1: 180-189
- nZaitsev, Yu. P. and Aleksandrov, B. G., (1997). Recent man-made changes in the Black Sea ecosystem. In: Sensitivity to Change: Black Sea, Baltic Sea and North Sea, E. Ozsoy and A. Mikaelyan, editors, Kluwer Acad. Publ., pp. 25-32
- nZaitsev, Yu. P. and Alexandrov, B. G., (1995). Recent man-made changes in the Black Sea ecosystem. Sensitivity of North Sea. Baltic Sea and Black Sea to antropogenic and climatic changes. Workshop, Varna- Bulgaria, 14-18 Nov
- nZhorov, V.A. and Boguslavsky, S.G., (1985). Tendencies of Some Hydrological- Hydrochemical Processes of the Black Sea, Meteorology and Hydrology 11: 63- 69.


