Keywords
Iron catalyzed oxidation, Carbonyl content, SDS-PAGE, Malonaldehyde, Grey mullet
Introduction
Lipid oxidation has detrimental effects on the nutritional and functional properties of fish and fish products accompanied by an offflavour or off-odour. Oxidizing of protein and their damages in the biological system has been studied over the past decades. However, the information about protein oxidation in fish muscle has not been given much attention (Kjærsgård and Jessen, 2004). There is no doubt that oxidative damages of proteins by hydroxyl radicals with subsequent alteration of their functions are unavoidable in aqueous phase in fish muscle (Srinivasan & Hultin, 1997; Soyer & Hultin, 2000). Limited studies on protein oxidation in fish muscle showed that the oxidation of protein can cause a decrease in protein solubility, an increase in protein carbonyl, and some loss in sulfhydryl groups and myosin heavy chain (Srinivasan & Hultin, 1997). These changes are known as protein denaturation and attributed to the denaturation/degradation of the structural muscle proteins (Saeed, Fawthrop & Howell, 1999; Soyer & Hultin, 2000; Saeed & Howell, 2004).
Iron that is important catalysts with their labile electron system is well suited to catalyze redox reactions (Kanner et al., 1988). The combination of H2O2 and a ferrous salt, which makes use of Fenton chemistry, is now considered to be the major mechanism by which the highly reactive hydroxyl radical that is generated in biological systems (Gutteridge et al., 1981; Kanner et al., 1986; Halliwell and Gutteridge, 1999). This mechanism is known as Harber-Weiss reaction (Kehrer, 2000) which is given below.
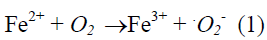
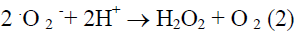
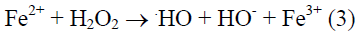
Hydroxy radical, a strong electrophilic compound, can easily react with the double bond of unsaturated lipids and propagate the chain reaction of lipid autoxidation (Choe and Min, 2006). It can also react with the side chains of amino acid residues at the metal binding side and cause oxidation of proteins in biological systems (Stadtman and Oliver, 1991).
Iron as a transition metal may be the most important lipid oxidation pro-oxidant in dark muscle fish due to its high reactivity and concentrations (Mei et al., 1998; Richards et al., 2002; Richards & Hultin, 2003; Undeland et al., 2004) and lipid oxidation in dark fish muscle increases in the presence of chelated iron or heme compounds (Hultin, 1992; Mei et al., 1998; Richards et al., 2002; Richards & Hultin, 2003; Undeland et al., 2004). Researchers have been suggesting that iron was the major catalyst of lipid oxidation in mullet dark muscle (Fisher and Deng, 1977) and liver (Ferreira et al., 2005). However, the effect of iron and hydroxyl radicals on oxidative damage of proteins in grey mullet is unknown. Many studies show that the Fe+2/H2O2 mixture is determined as the most efficient systems among the different metal catalyzed oxidation systems (Mercier et al., 2004; Martinaud et al., 1997; Nagasawa et al., 2001). From this point of view, Fe+2/H2O2 induced oxidation of lipid and proteins of mullet in vitro may be good indicator to determine oxidative damage. The determination of these changes is of great commercial importance for grey mullet, which has economic value in Turkey as fish and roe in order to determine the storage life. Therefore, the aim of this study was to evaluate the prooxidative effect of iron catalyzed oxidation in a Fenton-type (Fe+2SO4/H2O2) model system.
Material and Methods
The grey mullet (Mugil cephalus) was purchased from a local fish market in Adana (Turkey). The fish were transported to the laboratory in ice and then they were gutted, washed, filleted and homogenized. The samples were either processed for oxidation procedure immediately or frozen at -80 °C until they were analyzed.
Oxidation Procedure
Fish muscle (1g) was homogenized in 10 ml of 100 mM sodium phosphate buffer (pH 7.0) using a homogenizer (Ultra- turrax, Ika T8) for 1 min at high speed. Ferrous sulphate (0.5 mM) and hydrogen peroxide (1 mM) were added to homogenate. 1/2 Fe+2/H2O2 ratio was used to obtain maximum effect of oxidative system as recommended by Mercier et al (2004). The mixtures were incubated for 0, 3 and 5 h at 37 °C in a water bath, and the reaction was stopped by adding of butylated hydroxytolune (BHT, to 0.02% final concentration) to aliquots of 2 ml of homogenate. The mixture was either analyzed for lipid oxidation immediately or frozen at -80 °C until they were used.
Determination of Lipid Oxidation
Lipid oxidation was evaluated by production of thiobarbutiric acid reactive substances (TBARs) according to the method of Lynch & Frei (1993). 0.25 ml of 1 % (w/v) 2- thiobarbutiric acid in 0.05 M NaOH and 0.25 ml of 2.8 % (w/v) trichloroacetic acid were added to 0.5 ml of incubated samples. The samples were then incubated in a boiling water bath for 10 min, cooled and extracted with 2 ml of n-butanol. After clarifying the pink chromagen, the color intensity of n-butanol layer was measured at 535 nm against a blank of n-butanol. The concentrations of TBARs in samples were calculated by reference to a standard curve by using 1,1,3,3,- tetramethoxypropane. The results were expressed as mg malonaldehyde (MDA) per kg fish muscle.
The Determination of Protein Oxidation
Protein carbonyl content
The protein carbonyl contents were detected by reactivity with 2,4 dinitrophenylhydrazine (DNPH) as described by Levine, Williams, Stadtman & Shacter (1994). Two equal aliquots of incubated samples for 0, 1, 3 and 5 h were precipitated with 10% trichloracetic acid (w/v) and then centrifuged at 12 000 rpm for 3 min. One pellet was treated with 2 M HCl, and the other was treated with 10 mM DNPH in 2 M HCl. Samples were placed in a sample holder and shaken continuously for 1 h at room temperature. The two fractions were then precipitated with 10% TCA and centrifuged at 12 000 rpm for 3 min. The pellets were washed with 1 ml of ethanol: ethyl acetate (1:1 v/v) to remove free DNPH reagent. The samples were then centrifuged at 12,000 rpm for 3 min, and the supernatants were discarded. The washing procedure was repeated three times in total. The proteins were finally dissolved in 1 ml of 6 M guanidine- HCl in 20 mM potassium phosphate buffer (pH 2.3, adjusted with trifluoroacetic acid). All samples were then centrifuged to remove any insoluble material. The concentration of DNPH was determined at 360 nm against the blank of 1 ml guanidine solution, and molar absorbtion coefficient of 22,000 M-1/cm was used to quantify the levels of protein carbonyls. Protein concentration was determined at 280 nm of absorbance in the samples using BSA in 6 M guanidine as standard. The results were expressed as nmol carbonyl /mg protein.
Derivation of oxidized proteins with DNPH for electrophoresis
Incubated samples were diluted with an equal volume of 12% sodium dodecyl sulphate (SDS) to a final concentration of 6 % SDS. The protein concentrations of samples were analyzed with Lowry protein assay (Lowry et al., 1951). Labeling of oxidized protein was determined by Levine et al (1994). 20 mM DNPH in 10% (v/v) triflouracetic acid anhydride (TFA) was added in equal volume of incubated samples containing 6% SDS. The reaction was performed for 30 min in the dark and the stopped by addition of equal volume of incubated samples of neutralization solution containing 1.85 M Trisma-base, 28% glycerol, and 100 mM dithiothreitol (DTT) and then centrifuged at 12,000 rpm for 3 min. Sodium Dodecyl Sulphate-Poyacrylamide Gel Electrophoresis (SDS-PAGE) was used to oxidation induced polymerization of oxidized protein. SDS-PAGE was performed %10 resolving gel and %4 stacking gel according to method of Laemmli (1970) using BioRad minigel electrophoresis. Gels were stained with a solution of 0.025% Coomassie Blue R-250, 40% methanol and 7% acetic acid, and stained again with a solution of 5% methanol and 7% acetic acid.
Results and Discussion
Lipid Oxidation
The results of TBARs values (mg malonaldehyde/ kg fish muscle) of grey mullet muscle induced by fenton-like oxidation system are shown in Figure 1.
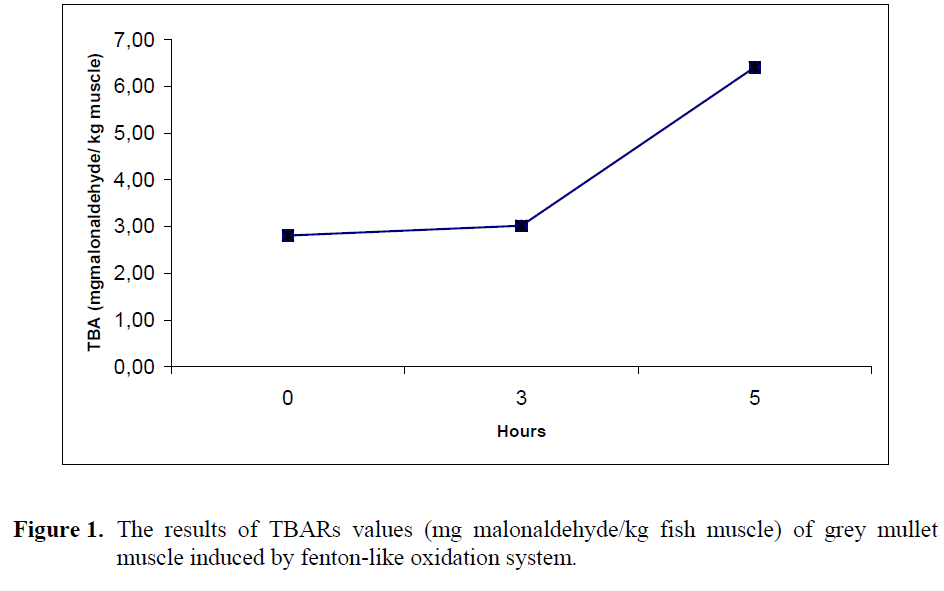
Figure 1: The results of TBARs values (mg malonaldehyde/ kg fish muscle) of grey mullet muscle induced by fenton-like oxidation system.
Treatment of grey mullet muscle with Fe+2SO4/H2O2 oxidation system for 0 h (2.81±0.16) and 3 h (3.02±0.60) at 37 °C had no significant effect on TBA values (p>0.05). However, after 5 h (6.41±0.55) incubation, there was a significant increase in TBARs values of grey mullet (p<0.05). The ferrous state of iron that is free to participate in Fenton chemistry produce hydroxide ion and a hydroxyl free radical which accelerate the lipid oxidation by breaking down hydrogen and lipid peroxides (Dunford, 1987). Soyer & Hultin (2000) observed that the TBARs values increased with an increase in iron concentration, when cod sarcoplasmic reticulum induced by non-enzymatic iron ascorbate oxidation system. Similar observation was found by Mei et al (1998) that increasing Fe+2 coming from FeSO4 concentration caused an increase of TBA formation in SDS emulsified salmon oil. These findings are agreed with our observations.
Protein Oxidation
The time course of protein oxidation in grey mullet muscle as measured by carbonyls content (nmol carbonyl/ mg protein) is shown in Figure 2.
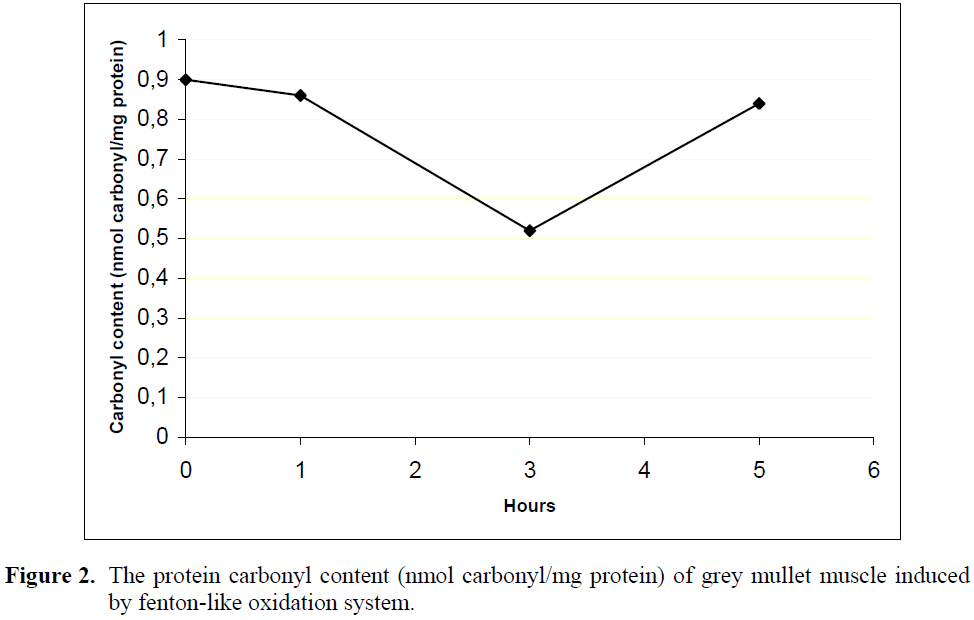
Figure 2: The protein carbonyl content (nmol carbonyl/mg protein) of grey mullet muscle induced by fenton-like oxidation system.
The initial carbonyl content in grey mullet muscle was 0.90±0.05 nmol carbonyl/mg protein. Although there was no significant change in carbonyl content within the first hour (0.86±0.07) of incubation with Fe+2SO4/H2O2 oxidation system, it significantly decreased to 0.52±0.01 nmol carbonyl/mg protein during 3 hours of incubation and then increased to 0.84±0.01 nmol carbonyl/ mg protein until 5 hour oxidation (p<0.05). Similarly, a decrease of carbonyl groups have been found by Mercier et al. (2004) in beef homogenate oxidized with Fe+2/H2O2 oxidation system with long incubation time. Such a decrease in carbonyl content has also been found in different oxidation systems (Batifoulier et al., 2002).
The coomassie staining of DNPH derivation of oxidized proteins
The coomassie staining of DNPH derivation of oxidized proteins in grey mullet is shown in Figure 3.
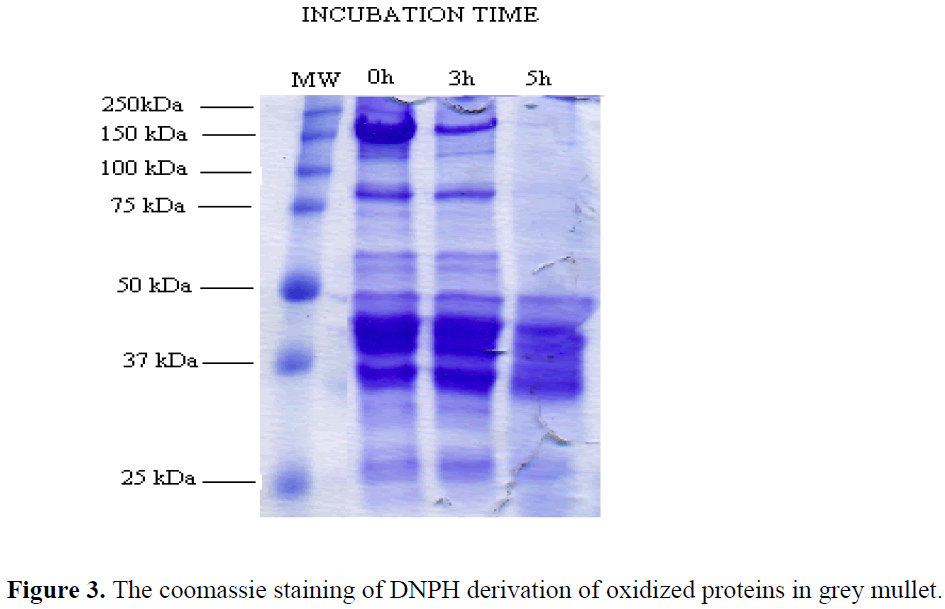
Figure 3: The coomassie staining of DNPH derivation of oxidized proteins in grey mullet.
The bands above 50 kDa disappeared in grey mullet after 5 h hour of incubation and the intensity of the bands near 100 kDa and 150 kDa decreased after 3 hour of incubation with Fe+2/H2O2 oxidation mixture in the presence of DTT (Fig1). Many studies have shown that the fragmentation and polymerization of oxidized proteins with metal catalyzed oxidation systems occur via disulfide cross-linking in the presence of reducing agent (Uchida et al., 1992; Liu & Xiong, 2000; Ooizumu and Xiong, 2004; Thanonkaew, Benjakul, Visessanguan & Decker, 2006). The dramatic changes in electronic pattern of grey mullet proteins, however, can be explained with forming of high molecular weight polymers through non-disulfide linkages in the presence of reducing agent. These bands can not be seen from the top of the gel in Figure 3 because they did not enter the gels. This result is supported by Decker et al (1993) which showed that nondisulfide covalent bonds can also cause high molecular weight polymerization for the reason that not all of the polymers were dissociable in the presence reducing agent.
Conclusion
As a result of this study, the iron catalyzed oxidation system was responsible for lipid oxidation and the unwanted changes electrophoretic patterns of grey mullet protein that the oxidation of proteins occurs nondisulfide covalent linking. These chemical modifications can alter the structure of a protein, usually leading to its degradation known as protein denaturation during processing and storage conditions. Until now, the relationships between aggregation, denaturation and loss of functionality of fish muscle proteins are only partially understood. Such studies about protein oxidation in fish species will be clearer to understand these irreversible changes in fish muscle in order to determine the storage life.
1761
References
- Batifoulier, F., Mercier, Y., Gatellier, P., Renerre, M. (2002). Infuence of vitamin E on lipid and protein oxidation induced by H2O2-activated MetMb in microsomal membranes from turkey muscle. Meat Science, 61: 389–395.
- Choe, E., Min, D.B. (2006). Chemistry and Reactions of Reactive Oxygen Species in Foods. Critical Reviews in Food Science and Nutrition, 46:1–22.
- Decker, E.A., Xiong, Y.L., Calvert, J.T., Crum, A.D., Blanchard, S.P.(1993). Chemical, physical, and functional properties of oxidized turkey white muscle myofibrillar proteins. J.Agric.Food Chem., 41: 186-189.
- Dunford, H.B. (1987). Free radicals in ironcontaining systems. Free Radical Biol. Med., 3: 405-421.
- Ferreira, M. Moradas-Ferreira, P., Reis- Henriques, M.A. (2005). Oxidative stress biomarkers in two resident species, mullet (Mugil cephalus) and flounder (Platichthys .esus), from a polluted site in River Douro Estuary. Portugal. Aquatic Toxicology, 71: 39–48
- Fisher, J., Deng, JC. (1977). Catalysis of lipid oxidation: A study of mullet (Mugil cephalus) dark flesh and emulsion model. J. Food Sci, 42:610.
- Gutteridge, J.M.C., Rowley, D.A., Halliwell, B. (1981). Superoxide dependent formation of hydroxyl radical in the presence of iron salts. Biochem. J., 199: 263-265
- Halliwell, B., Gutteridge, J.M.C. (1999). The chemistry of free radicals and related ‘reactive species’. Halliwell, B. & Gutteridge, J.M.C. (Eds.), Free Radicals in Biology and Medicine, 3rd edition , Oxford University Press: 36-100.
- Hultin, H.O. (1992). Biochemical deteriotion of fish muscle. Huss H.H., M. Jacobsen, J. Liston (eds.), Quality Assurance in fish industy, Elsevier Pub, Netherlands: 125- 139.
- Kanner, J., Harel, S., Hazan, B. (1986). Muscle Membranal Lipid Peroxidation by an “Iron Redox Cycle” System: Initiation by Oxy Radicals and Site-Specific Mechanism. J. Agric. Food Chem., 34: 506-510
- Kanner, J., Hazan, B., Doll, L. (1988). Catalytic “Free” Iron Ions in Muscle Foods. J. Agric. Food Chem. 36: 412-415.
- Kehrer, J.P. (2000). The Haber–Weiss reaction and mechanisms of toxicity. Toxicology, 149: 43–50.
- Kjærsgaard, I.V.H., Jessen, F., (2004). Oxidation of proteins in rainbow trout muscle. Presentation at the 34th WEFTA Annual Meeting, 12.-15. September, Lübeck, Germany
- Laemmli, U.K., (1970). Cleavage of stuctural proteins during the assembly of the head of bacteriophage T4. Nature, 277: 680-685.
- Levine, R.L., Williams, J.A., Stadtman, E.R., Shacter, E. (1994). Carbonyls assays for determination of oxidatively modified proteins. Meth. Enzymology, 233: 346- 357.
- Liu, G., Xiong, Y.L. (2000). Electrophoretic pattern, thermal denaturation, and in vitro digestibility of oxidized myosin. J. Agric. Food Chem., 48: 624-630
- Lowry, O.H., Rosebrough, N.J., Farr, A.L., Randall, R.J. (1951). Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
- Lynch, S.M., Frei, B. (1993). Mechanism of copper- and –iron- dependent oxidative modification of human low density lipoprotein. J. Lipid Research, 34: 1745- 1753.
- Martinaud, A., Mercier, Y., Marinova, P., Tassy, C., Gatellier, P., Renerre, M. (1997). Comparison of oxidative processes on myofibrillar proteins from beef during maturation and by different model oxidation systems. J. Agric. Food Chem., 45: 2481-2487
- Mei, L., Decker, E.A., McClements, D.J. (1998). Evidence of iron association with emusion droplets and its impact on lipid oxidation. J. Agric. Food Chem., 46: 5072- 5077.
- Mercier, Y., Gateller, P., Renerre, M. (2004). Lipid and protein oxidation in vitro, and antioxidant potential in meat from Charolais cows finished on pasture or mixed diet. Meat Science, 66(2): 467–473.
- Nagasawa, T., Yonekura, T., Nishizawa, N., Kitts, D.D. (2001). In vitro and in vivo inhibition of muscle lipid and protein oxidation by carnosine. Mol Cellular Biochem., 225: 29–34.
- Okizumi, T., Xiong, Y.L. (2004). Biochemical susceptibility of myosin in chicken myofibrils subjected to hydroxyl radical oxidizing systems. J. Agric. Food Chem., 52: 4303-4307.
- Richards, M.P., Hultin, H.O. (2003). Hemolysates from mackerel, herring and trout promoto lipid oxidation at different rates. Fish Sci., 69: 1298-1300.
- Richards, M.P., Østdal, H., Andersen, H.J. (2002). Deoxyhemoglobin-mediated lipid oxidation in washed fish muscle. J. Agric. Food Chem., 50: 1278-1283.
- Saeed, S., & Howell, N.K. (2004). Rheological and differential scanning calorimetry studies on structural and textural changes in frozen Atlantic mackerel (Scomber scombrus). J. Sci. Food Agric., 84: 1216- 1222.
- Saeed, S., Fawthrop, S.A., Howell, N.K. (1999). Electron spin resonance (ESR) study on free radical transfer in fishprotein interaction. J. Sci. Food Agric., 79:1809-1816.
- Soyer, A., Hultin, H.O. (2000). Kinetics of oxidation of the lipids and proteins of cod sarcoplasmic reticulum. J. Agric. Food Chem., 48: 2127-2134
- Srinivasan, S., Hultin, H.O. (1997). Chemical, Pyhsical, and Functional properties of cod proteins modified by a nonenzymic freeradical- generating system. J. Agric. Food Chem., 45: 310-320.
- Stadtman, E.R., Oliver, C.N. (1991). Metalcatalyzed oxidation of proteins. J. Bio. Chem., 266: 2005-2008.
- Thanonkaew, A., Benjakul, S., Visessanguan, W., Decker, E.A. (2006). The effect of metal ions on lipid oxidation, color and physicochemical properties of cuttlefish (Sepia pharaonis) sunjected to multiple freze-thaw cycles. Food Chem., 95: 591- 599.
- Uchida, K., Stadtman, E.R. (1992). Modification of histidine residues in proteins by reaction with 4- hydroxynonenal. Proc Natl Acad Sci, 89(10): 4544–4548.
- Undeland, I., Kristinsson, H.G., Hultin, H.O. (2004). Hemoglobin-mediated oxidation of washed minced cod muscle phospholipids:effect of pH and hemoglobin source. J. Agric. Food Chem., 52: 4444-4451.


